Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Tecniche per l’analisi della regolazione trascrizionale:
Analisi dei promotori (in vivo e in vitro) EMSA (SAGGIO DI MOBILITA’ ELETTROFORETICA) Footprint con DNAsi I Saggio di ipersensibilita’ alla DNAsi I Footprinting genomico (in vivo) Saggio di ImmunoPrecipitazione della Cromatina (ChIP) Analisi del posizionamento dei nucleosomi tramite nucleasi micrococcica Approcci per il clonaggio di fattori trascrizionali
 EMSA (SAGGIO DI MOBILITA’ ELETTROFORETICA) Footprint con DNAsi I. Saggio di ipersensibilita’ alla DNAsi I. Footprinting genomico (in vivo) Saggio di ImmunoPrecipitazione della Cromatina (ChIP) Analisi del posizionamento dei nucleosomi tramite nucleasi micrococcica. Approcci per il clonaggio di fattori trascrizionali.")
2
Analisi dei promotori: la trascrizione in vitro
Gli elementi di controllo della trascrizione sono clonati in un plasmide di fronte a una cassetta per la trascrizione Il plasmide linearizzato viene mescolato con estratti nucleari crudi trascrizionalmente attivi, o con frazioni e fattori purificati in presenza di NTPs + 32P GTP Il prodotto di trascrizione viene quantificato su gel denaturante di acrilamide Si analizza sia l’attivita’ trascrizionale sia l’accuratezza del sito di inizio

3
Analisi funzionale di sequenze regolatorie: i saggi reporter
(particolarmente utili in trasfezioni transienti) Geni reporter: CAT (cloramfenicolo-acetil-transferasi) Luc: (luciferasi) beta-galattosidasi GFP (green fluorescent protein)
 Geni reporter: CAT (cloramfenicolo-acetil-transferasi) Luc: (luciferasi) beta-galattosidasi. GFP (green fluorescent protein)")
4
Analisi funzionale di sequenze regolatorie: i saggi reporter
(particolarmente utili in trasfezioni transienti) Geni reporter: CAT (cloramfenicolo-acetil-transferasi) NB Controllo interno per efficienza di trasfezione, x es promotore forte con luciferasi promotore
 Geni reporter: CAT (cloramfenicolo-acetil-transferasi) NB Controllo interno per efficienza di trasfezione, x es promotore forte con luciferasi. promotore.")
5
Geni reporter: firefly luciferasi
NB Controllo interno per efficienza di trasfezione, x es promotore forte con renilla luciferasi

6
Geni reporter: GFP (Green Fluorescent Protein)
")
7
Sequenze regolatorie che si possono analizzate tramite i saggi reporter:
promotori enhancer (5’ e 3’) silencers elementi di controllo al 3’UTR attivita’ di microRNAs
 silencers. elementi di controllo al 3’UTR. attivita’ di microRNAs.")
8
Analisi di un promotore per delezioni successive

9
Generazione di mutanti per sostituzione (linker scanning)
")
10
Saggi di trasfezione stabile:
Saggi funzionali ancora piu’ stringenti: generazione di topi transgenici

11
Analisi funzionale degli elementi regolatori identificati sui promotori: analisi delle interazioni DNA-proteine

12
Il saggio EMSA (Electrophoretic Mobility Shift Assay) identifica interazioni tra DNA e proteine IN VITRO Mix: Estratto nucleare Buffer Competitore non specifico Incubare 10’ ice Add Probe (oligonucleotide ds marcato) Incubare 10’ RT Elettroforesi a freddo Disidratazione del Gel Esposizione/sviluppo
 Incubare 10’ RT. Elettroforesi a freddo. Disidratazione del Gel. Esposizione/sviluppo.")
13
Saggio EMSA: competizione, supershift

14
Il footprint con DNasi I permette di localizzare il sito di interazione tra proteina e DNA
Lodish Figure 10-6 NB: importanza di saturare i siti-estratti molto concentrati

15
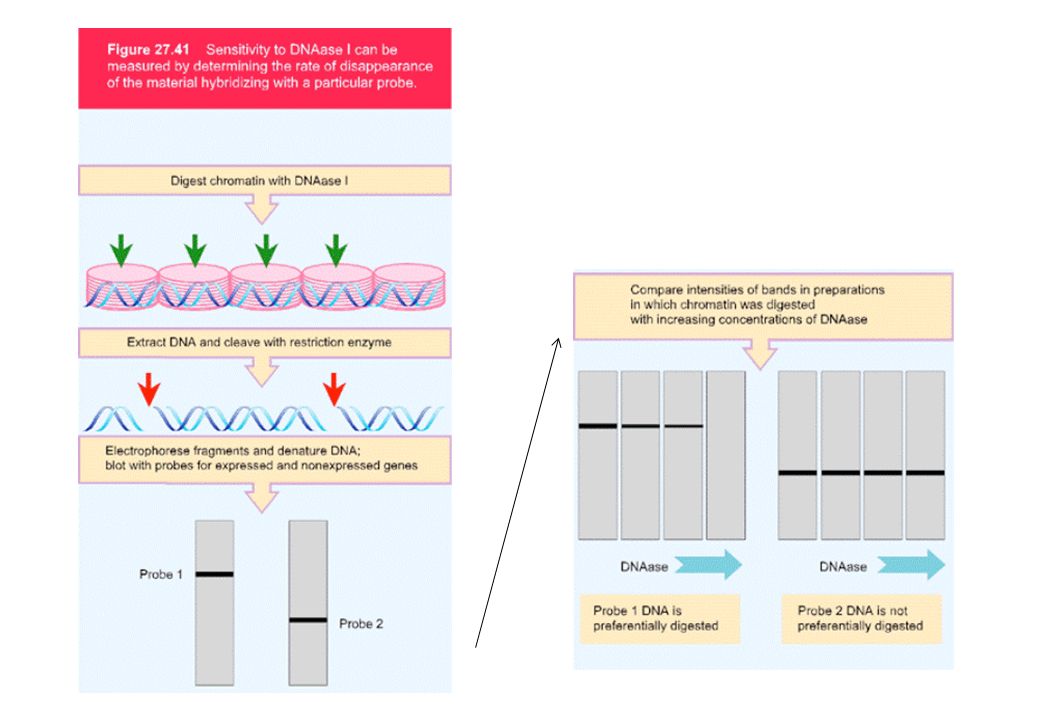
Saggio di ipersensibilita’ alla DNasi I
Regioni ipersensibili alla DNAsi I si trovano in connessione con geni attivamente trascritti: nelle regioni codificanti nelle regioni regolatrici Il saggio di ipersensibilita’ alla Dnasi I puo’ rivelare regioni regolatrici remote Le basi esatte per l’ipersensibilita’ non si conoscono (assenza di nucleosomi, distorsioni della doppia elica dovute a legame di fattori) Regioni di ipersensibilita’ alla Dnasi I sono state trovate in associazione a Promotori Enhancers LCRs silencers MARs (Matrix Attachment Regions o Scaffold-Associated Regions) insulators
 Regioni di ipersensibilita’ alla Dnasi I sono state trovate in associazione a. Promotori. Enhancers. LCRs. silencers. MARs (Matrix Attachment Regions o Scaffold-Associated Regions) insulators.")
17
Ligation-mediated PCR (alternativa al Southern genomico per mappare i siti ipersensibili):
:")
18
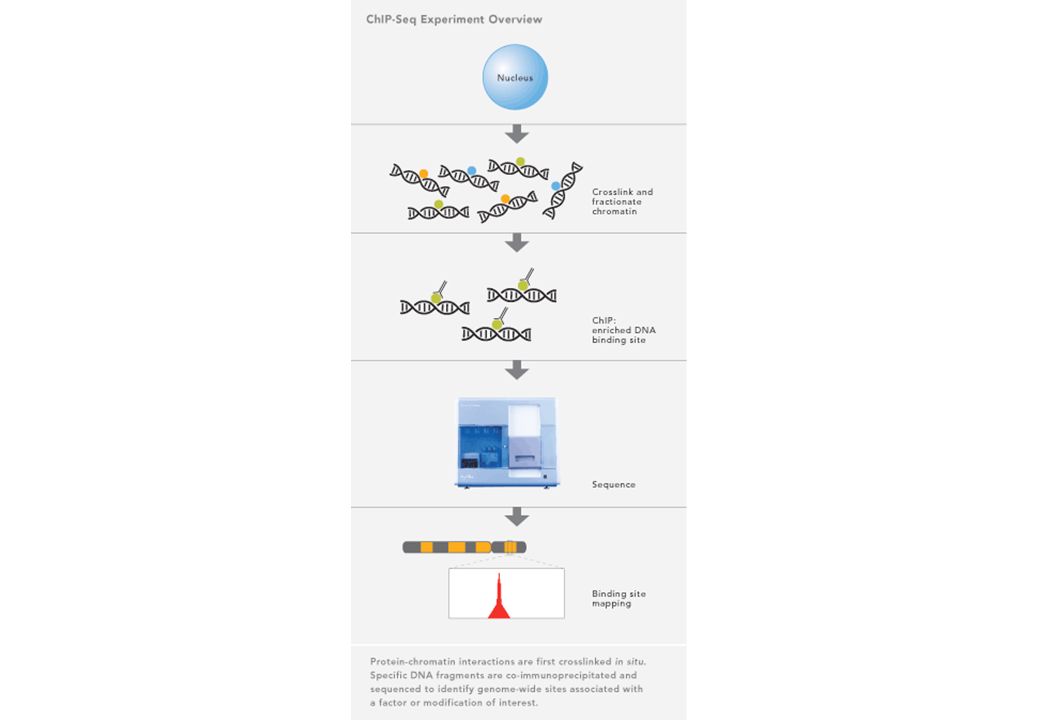
Identificare le proteine legate al DNA in vivo: saggio di immunoprecipitazione della cromatina (Chromatin immunoprecipitation-ChIP)
")
19
ChIP: - Crescere le cellule
- Cross-link con Formaldeide per legare covalentemente DNA e proteine Lisare e estrarre la cromatina Sonicare per frammentarla Immunoprecipitare i complessi DNA-proteina con Abs specifici Purificare il DNA degradando le proteine e revertire il cross-link Analizzare per la presenza di specifiche sequenze tramite PCR ChIP:

20
Le ChIP possono essere analizzate tramite real time PCR

21
Schiavone et al., Biochemical Journal 2009, 421, 283-292

22
La ChIP permette di analizzare anche specifiche modificazioni, per es
La ChIP permette di analizzare anche specifiche modificazioni, per es. sugli istoni (il codice istonico) Lodish Figure 10-59
 Lodish. Figure")
23
Il posizionamento dei nucleosomi sul DNA non e’ casuale e dipende:
dalla sequenza da proteine che legano il DNA e che possono influenzare il posizionamento da complessi proteici che rimodellano la cromatina Alcuni TF possono legare il DNA complessato nei nucleosomi, altri no e hanno quindi prima bisogno che i nucleosomi vengano riposizionati: rimodellamento

24
Posizionamento dei nucleosomi
Legame di fattori di trascrizione alla cromatina: Nelle regioni internucleosomiche Legame cooperativo dal bordo verso l’interno del nucl. in presenza di siti di legame multimerici Sequenza di riconoscimento selettivamente esposta verso l’esterno del nucl. Attivita’ ATPasi-dipendenti che rimodellano i nucleosomi vengono reclutate ai promotori e sono importanti per attivare la trascrizione

25
Tecniche per determinare la posizione dei nucleosomi
Digestione con la nucleasi micrococcica Saggio di accessibilita’ ad enzima di restrizione

26
Digestione con la nucleasi micrococcica

27
(A) MNase digestion of pBluescript assembled into nucleosomes
(A) MNase digestion of pBluescript assembled into nucleosomes. Lane 1:Marker; Lanes 2–6: MNase treatment for 0, 2, 4 and 8 minutes respectively. (B) MNase digestion of naked plasmid. Sen and De Benedetti BMC Molecular Biology :37 doi: /
 MNase digestion of pBluescript assembled into nucleosomes. Lane 1:Marker; Lanes 2–6: MNase treatment for 0, 2, 4 and 8 minutes respectively. (B) MNase digestion of naked plasmid. Sen and De Benedetti BMC Molecular Biology :37 doi: /")
28
Saggio di accessibilita’ ad enzima di restrizione
isolare i nuclei digerire con opportuno RE a concentrazioni crescenti purificare DNA genomico digerire con altro RE vicino oppure eseguire LM-PCR Analizzare tramite Southern blot con sonda opportuna oppure visualizzare i prodotti di PCR

29
by restriction enzyme accessibility assay and LM-PCR
Saggio di accessibilita’ ad enzima di restrizione Changes in chromatin structure of the TCRbeta locus in developing thymocytes by restriction enzyme accessibility assay and LM-PCR

30
Esempi di utilizzo di alcune di queste tecniche:
Agalioti et al. Ordered recruitment of chromatin modifying and general transcription factors to the IFNb promoter Cell 2000, 103,

31
Funzionamento dell’enhanceosoma: cascata di eventi:
Si assembla l’enhanceosoma Viene reclutato GCN5 che acetila i nucl. I e II (e HMG1, > interazioni con enhanc. e <il disassembl. dell’enhanc. mediato da CBP-dip. Acetilaz. di HMG1) 3) GCN5 si stacca, lasciando il posto all’ oloenzima CBP/PolII che a sua volta recluta: 4) SWI/SNF, che opera il rimodellamento del nucleosoma II (richiede acetilazione, prob. perché il bromodomain di BRG1 interagisce con gli H acetilati) A questo punto la TATA è libera e TBP può legare -> TRASCRIZIONE
 3) GCN5 si stacca, lasciando il posto all’ oloenzima CBP/PolII. che a sua volta recluta: 4) SWI/SNF, che opera il rimodellamento. del nucleosoma II (richiede acetilazione, prob. perché il bromodomain di BRG1. interagisce con gli H acetilati) A questo punto la TATA è libera e TBP. può legare -> TRASCRIZIONE.")
32
Figura 1a Micrococcal nuclease digestion
Determinazione della posizione dei nucleosomi Complete Mnase digestion + LM-PCR su cromatina cross-linkata Primers: La regione dell’enhancer è libera da nucleosomi (no PCR product -not shown) 2 nucleosomi (I e II) sono presenti a monte e a valle Il nucleosoma II è posizionato sul sito di inizio (da -15 a +132), solo 5 bp a valle della TATA box -170 +95 -227 +7 Figura 1a
 2 nucleosomi (I e II) sono presenti a monte e a valle. Il nucleosoma II è posizionato sul sito di inizio. (da -15 a +132), solo 5 bp a valle della TATA box Figura 1a.")
33
Figura 1b Assemblamento di nucleosomi in vitro
XbaI PvuII 275 198 326 B) Un frammento (-143/+183) che contiene la regione coperta dal nucleosoma II è in grado di assemblare un nucleosoma in vitro in posizione specifica (mentre il promotore di CMV no) Figura 1b
 Un frammento (-143/+183) che contiene la regione coperta. dal nucleosoma II è in grado di assemblare un nucleosoma. in vitro in posizione specifica (mentre il promotore di CMV no) Figura 1b.")
34
Assemblaggio in vitro dell’enhanceosoma su templato nucleosomale
EMSA Assemblaggio in vitro dell’enhanceosoma su templato nucleosomale EMSA con probe nucleosomale e fattori purificati I singoli fattori si possono legare da soli (accessibilità), come previsto dall’assenza di nucleosomi nella regione dell’enhancer Un complesso supermolecolare con minore mobilità si forma solo in presenza di HMG1 E non si forma se i 4 siti di legame per HMG1 sono mutati Figura 1
, come previsto dall’assenza di nucleosomi nella regione dell’enhancer. Un complesso supermolecolare con minore. mobilità si forma solo in presenza di HMG1. E non si forma se i 4 siti di legame per HMG1. sono mutati. Figura 1.")
35
Saggio di accessibilita’ a enzima di restrizione
L’infezione virale porta ad un rimodellamento del nucleosoma II, che diventa accessibile a NcoI. Si conferma che la regione dell’enhancer è libera da nucleosomi (accessibile a AvaII) B) L’acetilazione in vitro non è sufficiente a rimodellare il nucleosoma saggio di accessibilita’ all’Enz. Restr.: C) Però l’acetilazione aumenta la trascrizione in vitro (con HeLa NE) di 5 volte su templato nucleosomale. Sia con CBP che con GCN5 Forse l’acetilazione contribuisce a reclutare attività che rimodellino il nucleosoma al promotore dell’IFN Figura 2
 B) L’acetilazione in vitro non è sufficiente. a rimodellare il nucleosoma. saggio di accessibilita’ all’Enz. Restr.: C) Però l’acetilazione aumenta la trascrizione in vitro (con HeLa NE) di 5 volte su templato nucleosomale. Sia con CBP che con GCN5. Forse l’acetilazione contribuisce a reclutare. attività che rimodellino il nucleosoma. al promotore dell’IFN Figura 2.")
36
Figura 3 DNA-mediated pool down & Western Blot
L’acetilazione facilita il reclutamento di complessi che rimodellano la cromatina? Templato biotinilato Nucleosome assembly - dynabeads +/- GCN5 Enhanc. Assembly (HeLa NE +/- CBP) Western blot: reclutamento di BRG-1 e CBP L’enhanceosoma recluta sia la componente BRG1 di SWI/SNF, che CBP (HATs) Il reclutamento di BRG1 > 5x dopo acetilazione da parte di GCN5 mentre il reclutamento di CBP no CBP è però necessario per il reclutamento di BRG1 (reclutamento ad opera di CBP/oloenzima, non dell’enhanceosoma Figura 3
 Western blot: reclutamento di BRG-1 e CBP. L’enhanceosoma recluta sia la componente BRG1 di SWI/SNF, che CBP (HATs) Il reclutamento di BRG1 > 5x dopo acetilazione da parte di GCN5. mentre il reclutamento di CBP no. CBP è però necessario per il reclutamento di BRG1. (reclutamento ad opera di CBP/oloenzima, non dell’enhanceosoma. Figura 3.")
37
Figura 3 DNA-mediated pool down & Western Blot
L’acetilazione facilita il reclutamento di complessi che rimodellano la cromatina Templato biotinilato Nucleosome assembly - dynabeads +/- GCN5 Enhanc. Assembly (HeLa NE +/- CBP) Western blot: reclutamento di BRG-1 e CBP B) L’enhanceosoma recluta BRG1 E induce remodelling (accessibilità a NcoI e ClaI) Figura 3
 Western blot: reclutamento di BRG-1 e CBP. B) L’enhanceosoma recluta BRG1 E induce remodelling (accessibilità a NcoI e ClaI) Figura 3.")
38
L’enhancer è libero da nucleosomi, sia in vitro che in vivo
Il nucleosoma II, collocato sopra l’inizio di trascrizione, viene rimodellato (rimosso) in seguito a infezione virale (la condizione che attiva il promotore) L’acetilazione degli istoni in vitro (tramite GCN5 o CBP ricombinanti) NON è sufficiente per rimodellare il nucleosoma, però può aumentare l’attività trascrizionale in vitro DI UN TEMPLATO NUCLEOSOMICO in presenza di NE: l’acetilazione promuove il rimodellamento reclutando complessi che rimodellano la cromatina
 in seguito a infezione virale (la condizione che attiva il promotore) L’acetilazione degli istoni in vitro (tramite GCN5 o CBP ricombinanti) NON è sufficiente per rimodellare il nucleosoma, però può aumentare l’attività trascrizionale in vitro DI UN TEMPLATO NUCLEOSOMICO in presenza di NE: l’acetilazione promuove il rimodellamento reclutando complessi che rimodellano la cromatina.")
39
Chromatin Immunoprecipitation
ChIP: il reclutamento delle diverse componenti al promotore segue un ordine specifico: A) NFkB è il 1o fattore (picco 2h post-infezione): l’mRNA comincia ad essere espresso a 6h! B) Gcn5 è reclutata presto e declina in fretta, seguita da acetilazione di H4 (che correla con RNA) CBP: la cinetica di reclutamento non correla con il picco di acetilazione degli istoni -> Gcn5 è la principale attività HAT coinvolta PolII = CBP BRG1 dopo CBP, transiente: comincia il rimodellamento, non è richiesto per mantenerlo C) ChIP mRNA by RT-PCR D) TBP e TAF250 reclutati solo dopo il rimodellamento (6h) -> il loro reclutamento Correla finalmente con la trascrizione
 NFkB è il 1o fattore (picco 2h post-infezione): l’mRNA comincia ad essere espresso a 6h! B) Gcn5 è reclutata presto e declina in fretta, seguita da acetilazione di H4 (che correla con RNA) CBP: la cinetica di reclutamento non correla con il picco di acetilazione degli istoni -> Gcn5 è la principale attività HAT coinvolta. PolII = CBP. BRG1 dopo CBP, transiente: comincia il rimodellamento, non è richiesto per mantenerlo. C) ChIP. mRNA by RT-PCR. D) TBP e TAF250 reclutati. solo dopo il rimodellamento. (6h) -> il loro reclutamento. Correla finalmente con la. trascrizione.")
40
Funzionamento dell’enhanceosoma: cascata di eventi:
Si assembla l’enhanceosoma Viene reclutato GCN5 che acetila i nucl. I e II (e HMG1, > interazioni con enhanc. e <il disassembl. dell’enhanc. mediato da CBP-dip. Acetilaz. di HMG1) 3) GCN5 si stacca, lasciando il posto all’ oloenzima CBP/PolII che a sua volta recluta: 4) SWI/SNF, che opera il rimodellamento del nucleosoma II (richiede acetilazione, prob. perché il bromodomain di BRG1 interagisce con gli H acetilati) A questo punto la TATA è libera e TBP può legare -> TRASCRIZIONE
 3) GCN5 si stacca, lasciando il posto all’ oloenzima CBP/PolII. che a sua volta recluta: 4) SWI/SNF, che opera il rimodellamento. del nucleosoma II (richiede acetilazione, prob. perché il bromodomain di BRG1. interagisce con gli H acetilati) A questo punto la TATA è libera e TBP. può legare -> TRASCRIZIONE.")
41
Differenza tra enhanceosoma (IFN) e promotori/enhancer artificiali
ENHANCEOSOMA: assembla una superficie unica, formata dall’interazione dei diversi domini di attivazione che interagiscono a punti molteplici con CBP -> si raggiunge alta affinità e specificità che permettono il reclutamento efficiente dell’oloenzima CBP/PolII (poco abbondante) ->per IFN, il reclutamento di CBP coincide con il reclutamento di SWI/SNF, che induce il rimodellamento del nucl. II permettendo il recl. di TBP e TAF250 (TFIID). Il contrario (SWI/SNF reclutato prima di acetilazione) avviene nel caso del promotore HO di lievito ->specificita’/unicita’ di meccanismo ENHANCER SINTETICO: superficie di attivazione uniforme con solo un tipo di interazione con CBP -> complesso a bassa affinità e specificità, che permette l’assemblamento funzionale del complesso di inzio solo se TFIID viene reclutato prima, stabilizzando le interazioni con il promotore e permettendo il reclutamento del resto dei GTF.
 ->per IFN, il reclutamento di CBP coincide con il reclutamento di SWI/SNF, che induce il rimodellamento del nucl. II permettendo il recl. di TBP e TAF250 (TFIID). Il contrario (SWI/SNF reclutato prima di acetilazione) avviene nel caso del promotore HO di lievito. ->specificita’/unicita’ di meccanismo. ENHANCER SINTETICO: superficie di attivazione uniforme con solo un tipo di interazione con CBP -> complesso a bassa affinità e specificità, che permette l’assemblamento funzionale del complesso di inzio solo se TFIID viene reclutato prima, stabilizzando le interazioni con il promotore e permettendo il reclutamento del resto dei GTF.")
42
Estensioni “genome-wide” dei metodi di analisi della cromatina

43
Estensione della ChIP ad un’analisi “genome-wide: la tecnica del ChIP on Chip
(fl. label by LMPCR)
")
44
ULTERIORE Estensione della ChIP ad un’analisi “genome-wide:
la tecnica del ChIP & Seq DEEP SEQUENCING

46
ChIP (PCR or qPCR) ChIP on CHIP ChIP SEQ
 ChIP on CHIP ChIP SEQ")
49
Youtube animation ChIP-Seq

50
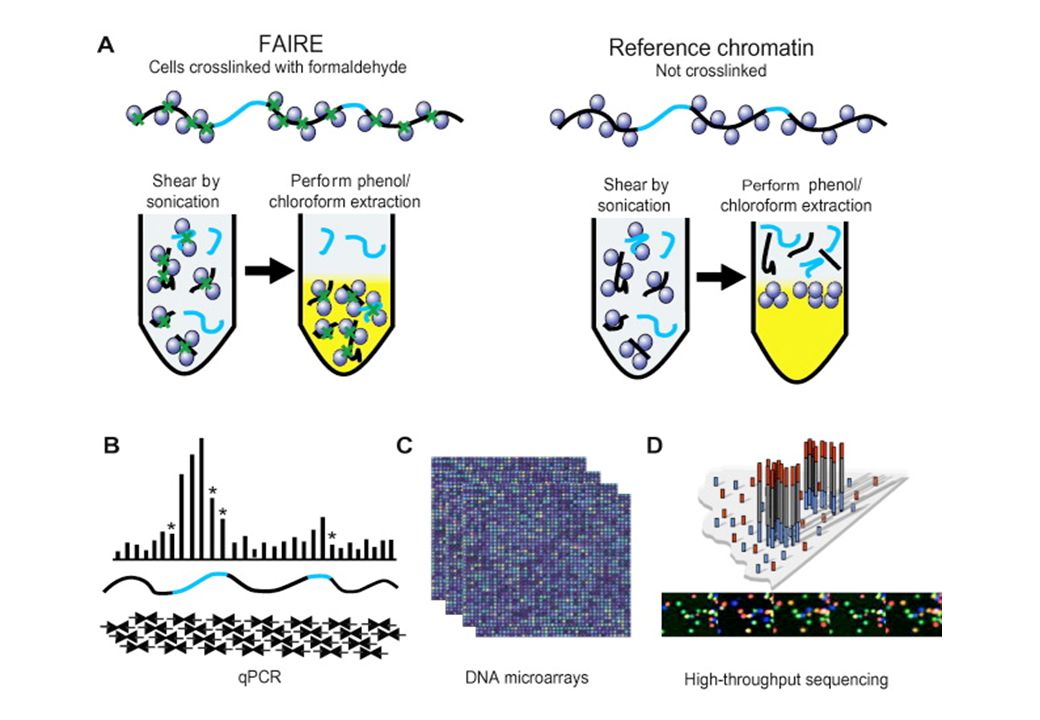
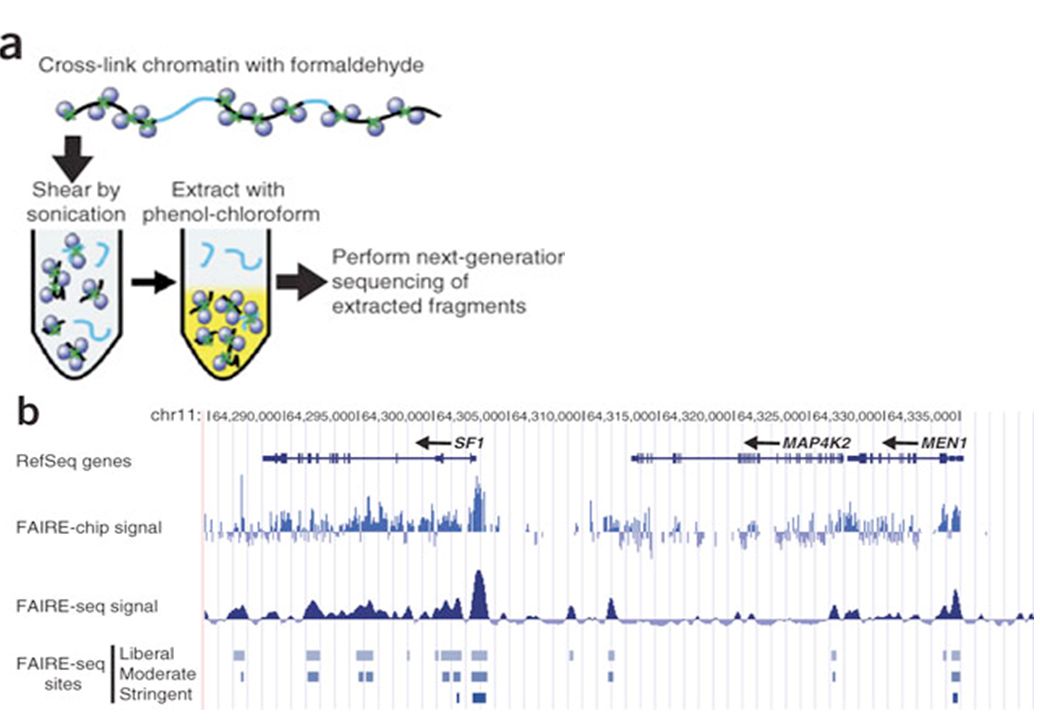
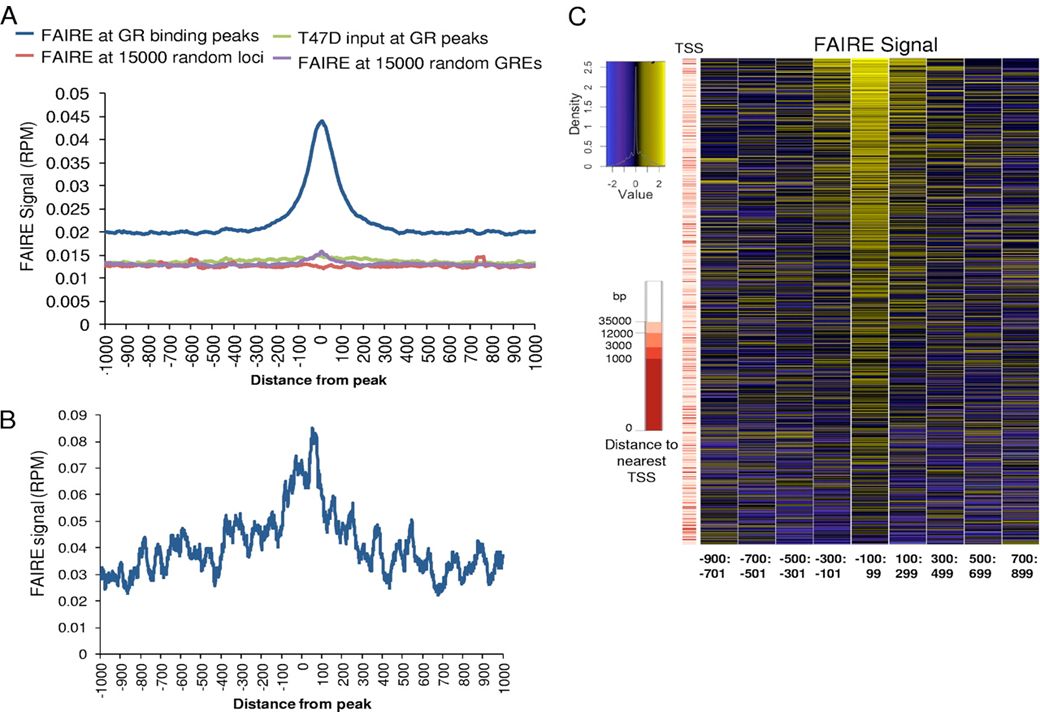
(Formaldehide-Assisted Isolation of Regulatory Elements)
Invece della DNAsi I per mappare regioni di cromatina libere da nucleosomi: FAIRE (Formaldehide-Assisted Isolation of Regulatory Elements)
")
54
Nucleosome-mapping technologies.
Nucleosome-mapping technologies. In vivo-isolated or in vitro-reconstituted chromatin is fractionated by micrococcal nuclease (MNase) digestion, which hydrolyzes the linker DNA in between nucleosomes, generating single nucleosomes. After proteinase treatment, ∼150-bp mononucleosomal DNA fragments are isolated by agarose gel electrophoresis. Subsequently, several techniques can be used to identify the collected DNA fragments and measure which fragments are enriched. Microarray hybridization and high-throughput sequencing allow the examination of nucleosome occupancy and positions on a genomic scale. Enriched fragments indicate that these sequences were integrated into a nucleosome, whereas depleted fragments were present in linker regions. See the text for details. Jansen A , and Verstrepen K J Microbiol. Mol. Biol. Rev. 2011;75:
 digestion, which hydrolyzes the linker DNA in between nucleosomes, generating single nucleosomes. After proteinase treatment, ∼150-bp mononucleosomal DNA fragments are isolated by agarose gel electrophoresis. Subsequently, several techniques can be used to identify the collected DNA fragments and measure which fragments are enriched. Microarray hybridization and high-throughput sequencing allow the examination of nucleosome occupancy and positions on a genomic scale. Enriched fragments indicate that these sequences were integrated into a nucleosome, whereas depleted fragments were present in linker regions. See the text for details. Jansen A , and Verstrepen K J Microbiol. Mol. Biol. Rev. 2011;75:")
55
cis and trans factors affecting nucleosome positioning in S. cerevisiae.
cis and trans factors affecting nucleosome positioning in S. cerevisiae. (A) The most important cis factor affecting local nucleosome positioning is the AT content of the local DNA. Poly(dA:dT) tracts act as antinucleosomal “barriers.” These barriers do not form a nucleosome, but highly positioned nucleosomes are often formed immediately adjacent to the barrier element. Second, the formation of nucleosomes is further enhanced by DNA sequences containing regularly spaced A/T dinucleotides (approximately one dinucleotide every 10 bp) and G/C dinucleotides (in between the A/T dinucleotides). Third, nucleosomes are also positioned by the steric hindrance of neighboring nucleosomes, much like beads on a string. This positioning signal is not perfect and deteriorates with increasing distance from strongly positioned nucleosomes, explaining why nucleosome positioning becomes increasingly “fuzzy” with increasing distance to positioning signals. (B) trans factors like ATP-consuming chromatin remodeling factors can remove (left) or slide (middle) nucleosomes. In addition, histone-modifying enzymes can add or remove covalent modifications to certain histone residues. For example, histone acetyltransferases (HATs) can add an acetyl residue, which can be removed by histone deacetylases (HDACs). Acetyl groups add an electronegative charge to the histones, which repulses the negatively charged DNA polymer, resulting in a modified, “looser” DNA-histone interaction and an increased accessibility of the DNA. Jansen A , and Verstrepen K J Microbiol. Mol. Biol. Rev. 2011;75:
 The most important cis factor affecting local nucleosome positioning is the AT content of the local DNA. Poly(dA:dT) tracts act as antinucleosomal barriers. These barriers do not form a nucleosome, but highly positioned nucleosomes are often formed immediately adjacent to the barrier element. Second, the formation of nucleosomes is further enhanced by DNA sequences containing regularly spaced A/T dinucleotides (approximately one dinucleotide every 10 bp) and G/C dinucleotides (in between the A/T dinucleotides). Third, nucleosomes are also positioned by the steric hindrance of neighboring nucleosomes, much like beads on a string. This positioning signal is not perfect and deteriorates with increasing distance from strongly positioned nucleosomes, explaining why nucleosome positioning becomes increasingly fuzzy with increasing distance to positioning signals. (B) trans factors like ATP-consuming chromatin remodeling factors can remove (left) or slide (middle) nucleosomes. In addition, histone-modifying enzymes can add or remove covalent modifications to certain histone residues. For example, histone acetyltransferases (HATs) can add an acetyl residue, which can be removed by histone deacetylases (HDACs). Acetyl groups add an electronegative charge to the histones, which repulses the negatively charged DNA polymer, resulting in a modified, looser DNA-histone interaction and an increased accessibility of the DNA. Jansen A , and Verstrepen K J Microbiol. Mol. Biol. Rev. 2011;75:")
56
Pugh Nat Str and Mol biol 2010
The upper panel shows a cross-section of a nucleosome, in which occupancy is distinguished from positioning. The lower panel shows how the two are measured. Occupancy is the area under the curve and reflects the local density of nucleosomes in a population, as illustrated by the column of spheres. Positioning or fuzziness is reflected in the standard deviation of the curve and is illustrated by how well the spheres are aligned in a column. The position of a nucleosome relative to some standard is indicated by how closely two peaks are separated. Comparing peaks of curves having high standard deviations is not likely to be meaningful because both peak locations have very high uncertainty.

57
Tecniche per studiare l’organizzazione spaziale del genoma:
FISH (localizzazione e co-localizzazione di DNA-RNA, RNA-proteina) Chromosome conformation capture (3C)
 Chromosome conformation capture (3C)")
58
superficie dei cromosomi X)
Nel nucleo interfasico, i cromosomi occupano spazi discreti, i territori cromosomali, separati da canali detti domini intercromosomali. I geni attivi sono preferenzialmente localizzati alla periferia dei territori, e gli RNA trascritti si formano alla superficie e vengono “riversati” nei domini intercr. Riduzione del volume a disposizione -> facilitazione della formazione dei complessi macromolecolari coinvolti in trascrizione e splicing. I cromosomi piu’ attivi trascrizionalmente avranno piu’ superficie a contatto con i canali (confermato dall’analisi della superficie dei cromosomi X) Le regioni attive e inattive dei diversi cromosomi appaiono esssere segregate in punti separati dei territori, suggerendo che la fibra di cromatina sia fortemente ripiegata, con continue anse tra la periferia e l’interno del nucleo. Studi di FRAP (fluorescence recovery after photobleaching) suggeriscono che particelle relativamente grandi di destrano e ficoll si muovono liberamente all’interno dei canali intercr., che sarebbero liberi da matrice di filamenti strutturali e fungerebbero da canali preferenziali che culminano nei pori nucleari e veicolano gli RNA verso l’apparato di esporto Sia i centromeri che le braccia dei cromosomi si muovono durante l’interfase, spesso in relazione al ciclo cellulare -> replicazione? Metafase interfase FISH del cromosoma 4 Structure and function in the nucleus Lamond and Earnshaw, 1998, Science 280, 547
 Le regioni attive e inattive dei diversi cromosomi appaiono esssere segregate in punti separati dei territori, suggerendo. che la fibra di cromatina sia fortemente ripiegata, con continue anse tra la periferia e l’interno del nucleo. Studi di FRAP (fluorescence recovery after photobleaching) suggeriscono che particelle relativamente grandi di. destrano e ficoll si muovono liberamente all’interno dei canali intercr., che sarebbero liberi da matrice di filamenti. strutturali e fungerebbero da canali preferenziali che culminano nei pori nucleari e veicolano gli RNA verso l’apparato. di esporto. Sia i centromeri che le braccia dei cromosomi si muovono durante l’interfase, spesso in relazione al ciclo cellulare -> replicazione Metafase interfase. FISH del cromosoma 4. Structure and function in the nucleus. Lamond and Earnshaw, 1998, Science 280, 547.")
59
attivi localizzano altrove.
Il silenziamento genico da parte dell’eterocromatina puo’ coinvolgere il movimento del gene silenziato da un’area all’altra del nucleo, probabilmente mediato da proteine come nel caso di Ikaros Ikaros e’ un regolatore trascrizionale essenziale per lo sviluppo linfoide. Di 6 geni inattivi o attivi in cellule pre-B o in cellule B mature, i geni inattivi co-localizzano sempre con Ikaros a siti di eterocromatina centromerica, mentre quelli attivi localizzano altrove. CD4 attivo inattivo Structure and function in the nucleus Lamond and Earnshaw, 1998, Science 280, 547

60
Chromosome Conformation Capture on Chip (4C)
Chromosome Conformation Capture & Seq (HiC) Chromosome Conformation Capture (3C)
 Chromosome Conformation Capture (3C)")
61
Interazioni in cis- e in trans- nelle transcription factories
Esperimenti di FISH, 4C e HiC confermano la co-localizzazione nucleare di alleli attivi di geni separati da decine di Mb in cis e anche in trans (cioe’ su diversi cromosomi): CO-REGOLAZIONE Fraser and Bickmore, Nature 2007,
: CO-REGOLAZIONE. Fraser and Bickmore, Nature 2007,")
Presentazioni simili