Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Antigeni-Antigenicità Immunogenicità

2
Antigeni (Ag) Tutte le molecole in grado di attivare il sistema immunitario sono detti antigeni Normalmente gli Ag sono sostanze estranee all’organismo ad alto peso molecolare quali proteine e lipopolisaccaridi Non sono antigeni molecole a basso peso molecolare, anche se estranee, come per esempio alcuni farmaci disaccaridi ecc… Essi si possono trovare sulla superficie delle cellule oppure si possono trovare liberi (antigeni circolanti) La capacità di un Ag di combinarsi con un Ab riflette la sua antigenicità
 La capacità di un Ag di combinarsi con un Ab riflette la sua antigenicità.")
3
Immunogenicità è la capacità di un Ag di indurre una risposta immune
Un Ag può essere antigenico ma non immunogeno, cioè si lega all’Ab ma non induce la produzione di Ab da parte della cellula B 1-5. AGENTI CAPACI DI INTERAGIRE CON IL SISTEMA IMMUNITARIO Antigeni ed immunogeni Una definizione esauriente degli antigene è quella di molecole capaci di interagire con il sistema immunitario. In tale concetto si possono distinguere due aspetti: 1. la immunogenicità, cioè la proprietà di indurre una risposta immune; 2. la antigenicità, cioè la capacità di essere riconosciute delle strutture specifiche della risposta immune quali i recettori di membrana dei linfociti T e B e gli anticorpi solubili. Ciò significa che tutte le molecole immunogeniche sono anche antigeniche, ma non tutti gli antigeni sono immunogeni. Dal punto di vista fisico-chimico gli immunogeni mostrano le seguenti caratteristiche: a) sono dotati di elevato peso molecolare e di notevole complessità chimica interna; b) sono solubili o facilmente solubilizzabili dalle cellule fagocitiche; c) sono in genere estranei alla specie in cui vengono introdotti. Il peso molecolare (p.m.) degli immunogeni è generalmente superiore a 10 kD. Proteine come 1'insulina (5,7 kD) e la protamina (6 kD) sono scarsamente immunogeniche. I polisaccaridi con p.m. anche superiore ai 10 kD (per esempio l'eparina che pesa 17 kD) sono poco immunogenici. Per contro, le grosse molecole proteiche (ribonucleasi, 14 kD; ovalbumina 40 kD; tossina tetanica, 55 kD; tireoglobulina 669 kD; emocianina, kD) sono tutti esempi di buoni antigeni. I destrani (tra 50 e 100 kD) sono anch'essi antigenici ma non per tutte le specie. All'interno di ciascuna molecola esistono regioni di dimensioni limitate che funzionano come determinanti antigenici od epitopi. Ciascuna di esse viene riconosciuta dal sistema immunitario distintamente dalle altre; il numero di questi siti aumenta ovviamente con l'aumentare delle dimensioni di una molecola. Le dimensioni delle molecole rappresentano comunque una condizione necessaria, ma non sufficiente, per la sua immunogenicità. È possibile produrre polimeri sintetici di qualsiasi peso, come il polistirolo, il nylon, il teflon, la poliacrilamide, gli omopolimeri degli aminoacidi, ma queste sostanze non risultano immunogeniche. Il motivo principale dell’assenza di antigenicità è la mancanza di complessità molecolare interna. Queste macromolecole hanno una struttura primaria relativamente semplice: uno o due monomeri legati covalentemente dentro una struttura ripetitiva che alla fine raggiunge un grande peso. Sia le proteine che i polisaccaridi naturali hanno invece una struttura chimica interna molto più complessa. Un'altra ragione della scarsa immunogenicità dei polimeri sintetici è la loro insolubilità nei fluidi corporei e la loro insensibilità alla solubilizzazione da parte degli enzimi. In pratica, tutte le molecole antigeniche devono essere solubili; gli stessi microrganismi vengono catturati dalle cellule fagocitiche e digeriti fino a rendere solubili i loro costituenti antigenici. Per essere antigenica una molecola deve provenire da una specie diversa o da un animale della stessa specie, ma completamente diverso da quello nel quale viene introdotta. Maggiore è la distanza o l'estraneità dalla specie da cui origina l'antigene, maggiore è l'entità della risposta immunologica dell'individuo ricevente.
 sono dotati di elevato peso molecolare e di notevole complessità chimica interna; b) sono solubili o facilmente solubilizzabili dalle cellule fagocitiche; c) sono in genere estranei alla specie in cui vengono introdotti. Il peso molecolare (p.m.) degli immunogeni è generalmente superiore a 10 kD. Proteine come 1 insulina (5,7 kD) e la protamina (6 kD) sono scarsamente immunogeniche. I polisaccaridi con p.m. anche superiore ai 10 kD (per esempio l eparina che pesa 17 kD) sono poco immunogenici. Per contro, le grosse molecole proteiche (ribonucleasi, 14 kD; ovalbumina 40 kD; tossina tetanica, 55 kD; tireoglobulina 669 kD; emocianina, kD) sono tutti esempi di buoni antigeni. I destrani (tra 50 e 100 kD) sono anch essi antigenici ma non per tutte le specie. All interno di ciascuna molecola esistono regioni di dimensioni limitate che funzionano come determinanti antigenici od epitopi. Ciascuna di esse viene riconosciuta dal sistema immunitario distintamente dalle altre; il numero di questi siti aumenta ovviamente con l aumentare delle dimensioni di una molecola. Le dimensioni delle molecole rappresentano comunque una condizione necessaria, ma non sufficiente, per la sua immunogenicità. È possibile produrre polimeri sintetici di qualsiasi peso, come il polistirolo, il nylon, il teflon, la poliacrilamide, gli omopolimeri degli aminoacidi, ma queste sostanze non risultano immunogeniche. Il motivo principale dell’assenza di antigenicità è la mancanza di complessità molecolare interna. Queste macromolecole hanno una struttura primaria relativamente semplice: uno o due monomeri legati covalentemente dentro una struttura ripetitiva che alla fine raggiunge un grande peso. Sia le proteine che i polisaccaridi naturali hanno invece una struttura chimica interna molto più complessa. Un altra ragione della scarsa immunogenicità dei polimeri sintetici è la loro insolubilità nei fluidi corporei e la loro insensibilità alla solubilizzazione da parte degli enzimi. In pratica, tutte le molecole antigeniche devono essere solubili; gli stessi microrganismi vengono catturati dalle cellule fagocitiche e digeriti fino a rendere solubili i loro costituenti antigenici. Per essere antigenica una molecola deve provenire da una specie diversa o da un animale della stessa specie, ma completamente diverso da quello nel quale viene introdotta. Maggiore è la distanza o l estraneità dalla specie da cui origina l antigene, maggiore è l entità della risposta immunologica dell individuo ricevente.")
4
Immunogenicità Fattori individuali
estraneità: il sistema immunitario è in grado di discriminare il “self” dal “non self”. Le molecole estranee ad un organismo hanno capacità immunogena peso molecolare: più è elevato il peso molecolare della molecola più sa possibile sarà una risposta immunogenica; complessità chimica e strutturale delle molecole: l’eterogenicità chimica causa immunogenicità. Ad es.: omopolimeri grandi hanno immunogenicità bassa rispetto ai polimeri dello stesso peso molecolare. Gli Ag proteici più immunogenici se più sono complessi (struttura terziaria e quaternaria) Densità epitopica: regioni di dimensioni limitate che funzionano come determinanti antigenici Solubilità o solubilizzazione da parte dei fagociti carica dell’Ag Fattori individuali 1-5. AGENTI CAPACI DI INTERAGIRE CON IL SISTEMA IMMUNITARIO Antigeni ed immunogeni Una definizione esauriente degli antigene è quella di molecole capaci di interagire con il sistema immunitario. In tale concetto si possono distinguere due aspetti: 1. la immunogenicità, cioè la proprietà di indurre una risposta immune; 2. la antigenicità, cioè la capacità di essere riconosciute delle strutture specifiche della risposta immune quali i recettori di membrana dei linfociti T e B e gli anticorpi solubili. Ciò significa che tutte le molecole immunogeniche sono anche antigeniche, ma non tutti gli antigeni sono immunogeni. Dal punto di vista fisico-chimico gli immunogeni mostrano le seguenti caratteristiche: a) sono dotati di elevato peso molecolare e di notevole complessità chimica interna; b) sono solubili o facilmente solubilizzabili dalle cellule fagocitiche; c) sono in genere estranei alla specie in cui vengono introdotti. Il peso molecolare (p.m.) degli immunogeni è generalmente superiore a 10 kD. Proteine come 1'insulina (5,7 kD) e la protamina (6 kD) sono scarsamente immunogeniche. I polisaccaridi con p.m. anche superiore ai 10 kD (per esempio l'eparina che pesa 17 kD) sono poco immunogenici. Per contro, le grosse molecole proteiche (ribonucleasi, 14 kD; ovalbumina 40 kD; tossina tetanica, 55 kD; tireoglobulina 669 kD; emocianina, kD) sono tutti esempi di buoni antigeni. I destrani (tra 50 e 100 kD) sono anch'essi antigenici ma non per tutte le specie. All'interno di ciascuna molecola esistono regioni di dimensioni limitate che funzionano come determinanti antigenici od epitopi. Ciascuna di esse viene riconosciuta dal sistema immunitario distintamente dalle altre; il numero di questi siti aumenta ovviamente con l'aumentare delle dimensioni di una molecola. Le dimensioni delle molecole rappresentano comunque una condizione necessaria, ma non sufficiente, per la sua immunogenicità. È possibile produrre polimeri sintetici di qualsiasi peso, come il polistirolo, il nylon, il teflon, la poliacrilamide, gli omopolimeri degli aminoacidi, ma queste sostanze non risultano immunogeniche. Il motivo principale dell’assenza di antigenicità è la mancanza di complessità molecolare interna. Queste macromolecole hanno una struttura primaria relativamente semplice: uno o due monomeri legati covalentemente dentro una struttura ripetitiva che alla fine raggiunge un grande peso. Sia le proteine che i polisaccaridi naturali hanno invece una struttura chimica interna molto più complessa. Un'altra ragione della scarsa immunogenicità dei polimeri sintetici è la loro insolubilità nei fluidi corporei e la loro insensibilità alla solubilizzazione da parte degli enzimi. In pratica, tutte le molecole antigeniche devono essere solubili; gli stessi microrganismi vengono catturati dalle cellule fagocitiche e digeriti fino a rendere solubili i loro costituenti antigenici. Per essere antigenica una molecola deve provenire da una specie diversa o da un animale della stessa specie, ma completamente diverso da quello nel quale viene introdotta. Maggiore è la distanza o l'estraneità dalla specie da cui origina l'antigene, maggiore è l'entità della risposta immunologica dell'individuo ricevente.
 Densità epitopica: regioni di dimensioni limitate che funzionano come determinanti antigenici. Solubilità o solubilizzazione da parte dei fagociti. carica dell’Ag. Fattori individuali AGENTI CAPACI DI INTERAGIRE CON IL SISTEMA IMMUNITARIO. Antigeni ed immunogeni. Una definizione esauriente degli antigene è quella di molecole capaci di interagire con il sistema immunitario. In tale concetto si possono distinguere due aspetti: 1. la immunogenicità, cioè la proprietà di indurre una risposta immune; 2. la antigenicità, cioè la capacità di essere riconosciute delle strutture specifiche della risposta immune quali i recettori di membrana dei linfociti T e B e gli anticorpi solubili. Ciò significa che tutte le molecole immunogeniche sono anche antigeniche, ma non tutti gli antigeni sono immunogeni. Dal punto di vista fisico-chimico gli immunogeni mostrano le seguenti caratteristiche: a) sono dotati di elevato peso molecolare e di notevole complessità chimica interna; b) sono solubili o facilmente solubilizzabili dalle cellule fagocitiche; c) sono in genere estranei alla specie in cui vengono introdotti. Il peso molecolare (p.m.) degli immunogeni è generalmente superiore a 10 kD. Proteine come 1 insulina (5,7 kD) e la protamina (6 kD) sono scarsamente immunogeniche. I polisaccaridi con p.m. anche superiore ai 10 kD (per esempio l eparina che pesa 17 kD) sono poco immunogenici. Per contro, le grosse molecole proteiche (ribonucleasi, 14 kD; ovalbumina 40 kD; tossina tetanica, 55 kD; tireoglobulina 669 kD; emocianina, kD) sono tutti esempi di buoni antigeni. I destrani (tra 50 e 100 kD) sono anch essi antigenici ma non per tutte le specie. All interno di ciascuna molecola esistono regioni di dimensioni limitate che funzionano come determinanti antigenici od epitopi. Ciascuna di esse viene riconosciuta dal sistema immunitario distintamente dalle altre; il numero di questi siti aumenta ovviamente con l aumentare delle dimensioni di una molecola. Le dimensioni delle molecole rappresentano comunque una condizione necessaria, ma non sufficiente, per la sua immunogenicità. È possibile produrre polimeri sintetici di qualsiasi peso, come il polistirolo, il nylon, il teflon, la poliacrilamide, gli omopolimeri degli aminoacidi, ma queste sostanze non risultano immunogeniche. Il motivo principale dell’assenza di antigenicità è la mancanza di complessità molecolare interna. Queste macromolecole hanno una struttura primaria relativamente semplice: uno o due monomeri legati covalentemente dentro una struttura ripetitiva che alla fine raggiunge un grande peso. Sia le proteine che i polisaccaridi naturali hanno invece una struttura chimica interna molto più complessa. Un altra ragione della scarsa immunogenicità dei polimeri sintetici è la loro insolubilità nei fluidi corporei e la loro insensibilità alla solubilizzazione da parte degli enzimi. In pratica, tutte le molecole antigeniche devono essere solubili; gli stessi microrganismi vengono catturati dalle cellule fagocitiche e digeriti fino a rendere solubili i loro costituenti antigenici. Per essere antigenica una molecola deve provenire da una specie diversa o da un animale della stessa specie, ma completamente diverso da quello nel quale viene introdotta. Maggiore è la distanza o l estraneità dalla specie da cui origina l antigene, maggiore è l entità della risposta immunologica dell individuo ricevente.")
5
Tutti gli Ag si legano agli Ab utilizzando piccole zone superficiali e specifiche dette determinati antigenici o epitopi. Una singola molecola di antigene può contenere diversi epitopi riconosciuti da anticorpi differenti.

6
Riconoscimento dell’Ag nei linf. B
Il riconoscimento dell’Ag da parte dei linfociti B comporta un legame diretto tra Ig e l’Ag intatto prendendo contatto con aa non contigui alla sequenza primaria, ma vengono accostati dal ripiegamento della proteina

7
Riconoscimento dell’Ag nei linf. T
Il TCR serve per il riconoscimento dell’Ag da parte dei linf T. I linf T sono stimolati da sequenze di aa contigui, talora nascoste all’interno della struttura nativa della proteina e non direttamente accessibili al recettore Pertanto è necessaria la processazione L’Ag viene riconoscito solo se legati al MHC

8
I linfociti T hanno una doppia specificità: per i residui di MHC e per l’antigene. Il recettore che
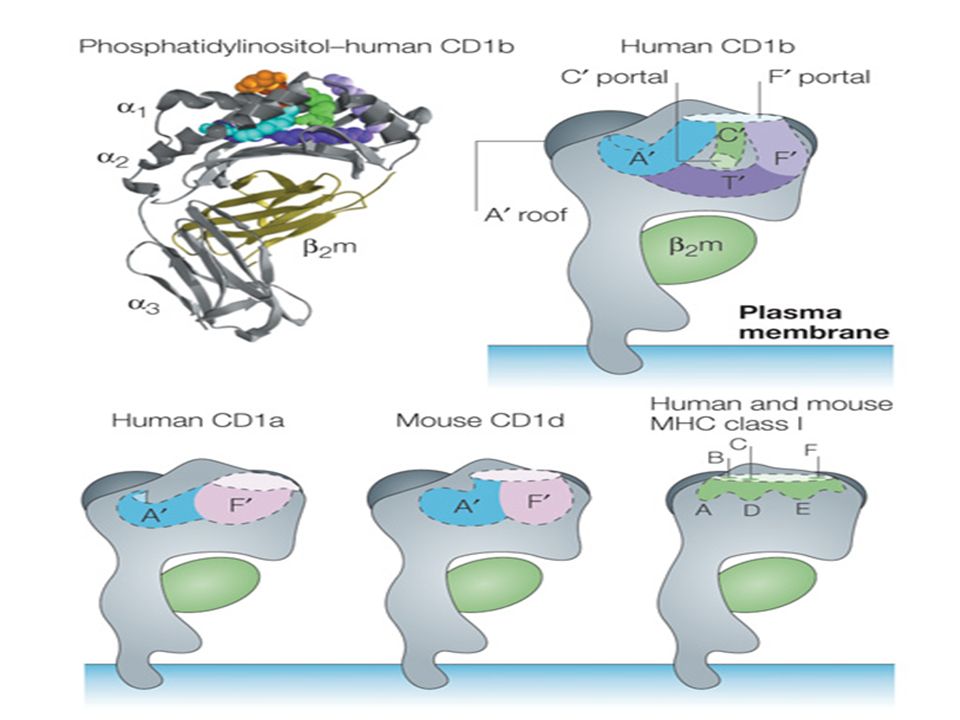
riconosce questi complessi MHC-antigene prende il nome di TCR (T-Cell Receptor) ed è distribuito in maniera clonale, cioè cloni di cellule T con diverse specificità avranno diversi TCR. Le cascate biochimiche di segnalazione iniziate dal TCR non sono da esso trasdotte: questo compito viene svolto dalle proteine invarianti CD3 e , che insieme al recettore formano il cosiddetto complesso TCR. La segnalazione è dunque legata a elementi estremamente variabili (il TCR) e a elementi costanti. Le cellule T esprimono altri recettori di membrana che non riconoscono l’antigene ma che contribuiscono alla risposta: in generale si parla di molecole accessorie. Il ruolo fisiologico è per alcune di facilitare la segnalazione del complesso TCR, per altre di fornire secondi segnali che attivano completamente le cellule. Altre molecole accessorie ancora servono a stabilizzare il legame con le APC, in modo da garantire il tempo necessario alla trasduzione del segnale. 1.1 -TCR per antigeni MHC-associati Il recettore antigenico sia dei linfociti CD4+ che dei CD8+ è un eterodimero costituito da una catena e una tra loro unite da un ponte disolfuro. Entrambe le catene sono costituite da un dominio simil-Ig N terminale variabile, uno simil Ig costante, un dominio transmembrana e una piccola regione citoplasmatica; la porzione extracellulare è dunque simile alla porzione legante l’antigene di un anticorpo, con una regione variabile e una costante sulla catena leggera e una variabile e una costante sulla catena pesante. Le regioni variabili V delle catene del TCR contengono piccole frazioni sulle quali la grande variabilità è concentrata: si parla di CDR, o Complementarity Determing Regions. Tre CDR sulla catena sono giustapposte a tre regioni simili sulla catena a formare la parte del recettore che riconosce i complessi peptide-MHC. La regione variabile della catena contiene poi una quarta regione ipervariabile che è il sito di legame per i cosiddetti superantigeni. Ogni catena del TCR è codificata da più segmenti genici che subiscono riarrangiamento somatico durante la maturazione dei linfociti. In entrambe le catene la terza regione ipervariabile è composta da sequenze codificate dai segmenti genici V e J (catena ) o dai segmenti V, D, J (catena ). Le regioni costanti C di entrambe le catene formano cerniere che contengono i residui di cisteina per i ponti disolfuro. I TCR e gli anticorpi sono strutturalmente simili, ma vi sono delle profonde differenze. I TCR non esistono in froma secreta e non hanno funzioni effettrici da soli, inoltre non subiscono variazioni nella regione C e non hanno maturazione della loro affinità. 1 1.1.1 Ruolo del TCR nel riconoscimento dell’antigene Il riconoscimento è mediato dalle regioni determinanti la complementarietà, o CDR, formate da entrambe le catene del TCR. Queste catene formano un singolo recettore eterodimerico responsabile sia della specificità per il peptide che per l’MHC. Il sito di legame per l’antigene è formato dalle sei CDR sulle due catene che formano un’interfaccia molto simile a quella degli anticorpi. Il contatto tra il TCR e il complesso peptide-MHC è limitato a uno o due aminoacidi: le cellule T riconoscono dunque i loro substrati sulla base di differenze minime. L’affinità del TCR per i complessi è bassa, molto più di quella della maggior parte degli anticorpi. Questa bassa affinità è probabilmente la ragione per cui sono necessarie le molecole di adesione per avere una risposta biologica. Il TCR e le sue molecole accessorie sul linfocita T si muovono in maniera coordinata ai loro ligandi sulle membrane delle APC per creare una struttura sopramolecolare detta sinapsi immunologica. 1.2 Proteine CD3 e del complesso TCR Le proteine CD3 e trasducono il segnale che porta all’attivazione del linfocita dietro stimolo del TCR. La molecola CD3 è in realtà un insieme di tre proteine designate , e ". Le tre proteine CD3 e la sono sempre uguali in tutte le cellule T: non hanno infatti ruolo nel riconoscimento ma solo nella trasduzione del segnale. 1.2.1 Struttura Le tre proteine CD3 sono omologhe tra loro, e le regioni extracellulari di tutte contengono un singolo dominio simil-Ig: queste tre proteine sono dunque membri della superfamiglia delle Ig. I domini citoplasmatici variano da 44 a 81 aminoacidi di lunghezza e ciascuno contiene una copia di una sequenza conservata detta ITAM (Immunoreceptor Tyrosine-based Activation Motif) che gioca un ruolo fondamentale nella segnalazione da parte del complesso. La catena ha una piccola regione extracellulare, una transmembrana e una lunga regione citoplasmatica contentente tre ITAM. L’espressione del complesso TCR richiede la sintesi di tutti i suoi componenti. Nei linfociti T maturi infatti l’intero complesso viene prodotto nel RE e trasportato sulla membrana. 2 1.2.2 Funzione Il primo evento intracellulare a seguito del riconoscimento antigenico è la fosforilazione dei residui di tirosina contenuti nei domini ITAM di CD3 e da parte di kinasi quali Lck o Fyn. Lck si associa alle code citoplasmatiche di CD4 e CD8, Fyn a CD3. Le fosfotirosine così create diventano siti di attacco per una tirosin kinasi, ZAP-70, che viene reclutata dalla catena e porterà alla variazione dell’espressione genica delle cellule T. 1.3 Recettori antigenici dei linfociti Il TCR è un secondo tipo di eterodimero espresso in un piccolo set di linfociti T -negativi; questo recettore è comunque associato alle proteine CD3 e . La maggioranza delle cellule con questo recettore non esprimono CD4 o CD8. Le cellule che esprimono questo recettore sono linearmente distinte da quelle che esprimono il più comune TCR: in totale meno del 5% dei linfociti esprime questa struttura. Queste cellule, insieme alle cellule NK-T, i linfociti B di tipo B-1 e le cellule MZ B, potrebbero rappresentare un importante collegamento tra l’immunità innata e quella adattativa. Le cellule con recettore non riconoscono gli antigeni peptidici MHC-associati e non sono MHC ristrette; alcune riconoscono piccole molecole fosforilate o lipidi comuni nei batteri, altre riconoscono proteine che non richiedono processamento o collaborazione delle APC. Il ruolo di queste cellule è comunque poco definito in quanto topi deficitari non si mostrano particolarmente immunodeficienti o maggiormente suscettibili alle infezioni batteriche. 1.4 Recettori antigenici delle cellule NK-T Una piccola popolazione di linfociti T esprime i markers tipici delle cellule NK: si tratta delle cellule NK-T. Le catene alfa del TCR di queste cellule hanno una diversità limitata e sono caratterizzate da un riarrangiamento caratteristico nell’uomo; queste cellule fanno ancora una volta da ponte tra l’immunità innata e quella adattativa. Tutte i TCR delle cellule NK-T riconoscono lipidi legati alle molecole simil-MHCI CD1. Queste cellule producono rapidamente citochine quali IL-4 e IFN- a seguito della stimolazione. 1.5 Corecettori e recettori costimolanti nelle cellule T I corecettori sono una categoria di proteine di membrana che amplificano il segnale di TCR; queste strutture legano le molecole di MHC. I costimolanti conducono anch’essi dei segnali che attivano le cellule T, ma riconoscono molecole sulle APC che non sono parte del complesso MHC-peptide. 1.5.1 CD4 e CD8: Corecettori coinvolti nell’attivazione delle cellule T MHC- ristrette Le cellule T mature esprimono CD4 o CD8, ma mai entrambi. Queste strutture interagiscono con entrambe le classi di MHC quando i TCR della cellula riconoscono i complessi MHC-peptide dell’APC. La funzione principale è nella trasduzione del segnale al momento del riconoscimento, ma possono 3 anche aumentare l’efficacia del legame tra cellula T ed APC. Nel pool dei linfociti maturi circa il 65% esprime CD4 e il 35% CD8. Struttura Entrambi i corecettori sono glicoproteine transmembrana facenti parte della superfamiglia Ig. CD4 viene espresso come monomero e presenta quattro domini simil-Ig extracellulari, una regione transmembrana e una coda basica citoplasmatica. I due domini simil-Ig amino terminali del CD4 legano il dominio 2 non polimorfico dell’MHCII. Le molecole CD8 esistono quasi sempre sotto forma di eterodimeri di due catene dette CD8 e CD8. Entrambe presentano un singolo dominio Ig extracellulare, una regione transmembrana e la coda citoplasmatica basica. Il dominio Ig di CD8 lega il dominio 2 non polimorfico delle molecole di MHCI. Funzione La separazione delle risposte dei linfociti CD4+ e CD8+ è dovuta alla capacità di queste molecole di legare solamente una classe di MHC e non l’altra. CD4 è in grado di legare MHCII e viene espresso sui linfociti i cui TCR riconoscono i complessi peptidici di questo tipo. Quasi tutti i linfociti CD4+ sono cellule di supporto che producono citochine. CD8 è in grado di legare le molecole MHCI. Quasi tutti i linfociti CD8+ sono linfociti citotossici il cui ruolo è sradicare le infezioni intracellulari. Esistono linfociti CD4+ con funzioni citotossiche ma sono comunque MHCII ristretti. CD4 e CD8 partecipano ai primi eventi segnalatori dopo il riconoscimento del complesso MHC-peptide. Queste funzioni sono mediate da una tirosin chinasi specifica dei linfociti T che prende il nome di Lck: questo enzima è associato in modo non covalente alle code sia del CD4 che del CD8. Quando un linfocita riconosce il complesso MHC l’interazione di CD4/8 con l’MHC porta il corecettore e la sua Lck nelle vicinanze del CTR; Lck a questo punto fosforila i domini ITAM delle proteine CD3 e e da il via alla segnalazione. 1.5.2 Recettori costimolanti ed inibitori della famiglia CD28 CD28 è una proteina che trasduce il segnale in associazione ai segnali in arrivo dal complesso TCR per attivare le cellule T naive. I linfociti T naive in generale necessitano di due segnali extracellulari distinti per proliferare e differenziare; il primo deriva dal legame dell’antigene al recettore e garantisce la specificità della risposta. Il secondo segnale viene fornito da molecole che vengono definite in generale costimolatori. I costimolatori meglio definiti per i linfociti T sono chiamati B7-1 (CD80) e B7-2 (CD86) e sono espressi sulle cellule dendritiche, sui macrofagi e sui linfociti B; queste molecole hanno specifici recettori sul linfocita. Il primo recettore per B7 scoperto fu la molecola CD28, espressa su quasi tutti i CD4+ e su metà dei CD8+. Il legarsi delle molecole B7 delle APC a CD28 fornisce al linfocita il segnale per esprimere proteine anti apoptosi, per produrre fattori di crescita e citochine e per promuovere proliferazione e differenziazione. 4 Un secondo recettore per molecole B7 venne successivamente scoperto e chiamato CTLA-4; questa struttura è omologa a CD28 e viene espressa sui linfociti recentemente attivati: la sua funzione è inibire l’attivazione controbilanciando i segnali in arrivo dal complesso TCR e da CD28. Molte altre strutture sono state scoperte in grado di legare le molecole B7 e sono equamente divise tra vie di attivazione e vie di terminazione. 1.5.3 CD2 e la famiglia SLAM di recettori costimolanti Un’importante famiglia di proteine che gioca un ruolo nell’attivazione delle cellule T ed NK è un gruppo di proteine strutturalmente legate ad un recettore detto CD2. Questo recettore contiene due domini Ig extracellulari, una regione di membrana e una lunga coda citoplasmatica. Nell’uomo il principale ligando è la molecola LFA-3 (Leukocyte Function-associated Antigen 3) che è espressa in molte cellule ematopoietiche e non. CD2 è esempio di molecola accessoria che funziona sia come una molecola di adesione che come trasduttore del segnale. Un sottogruppo distinto di protine CD2 è detto SLAM (Signaling Lymphocytic Activation Molecule). Le SLAM sono proteine integrali di membrana con due domini Ig extracellulari e una coda citoplasmatica che contiene un dominio detto ITSM (Immunoreceptor Tyrosin-based Switch Motif); il dominio si lega ad un adattatore detto SAP (SLAM Associated Protein) che contiene un dominio SH2 in grado di fare da ponte tra SLAM e Fyn, una chinasi. Un importante membro della famiglia SLAM è 2B4, mutazioni in questa molecola possono seriamente danneggiare il sistema immunitario. 1.5.4 Altre molecole accessorie dei linfociti T CD44 è una glicoproteina espressa in varie cellule, tra cui linfociti T maturi, timociti, cellule B, granulociti, macrofagi, eritrociti e fibroblasti. Questa molecola lega lo ialuronato e questa proprietà è responsabile della detenzione dei linfociti T nei siti extravascolari di infezione. Le cellule CD4+ attivate esprimono una proteina della famiglia del TNF detta CD40L, che lega CD40 dei linfociti B e di altre cellule e le attiva. CD40L è quindi un importante mediatore delle funzioni helper di questi linfociti. I linfociti attivati esprimono anche il FAS ligand; l’attivazione di FAS da parte di FASL porta all’apoptosi ed è importante per eliminare le cellule T iperstimolate. 5
 ed è distribuito. in maniera clonale, cioè cloni di cellule T con diverse specificità avranno diversi TCR. Le cascate. biochimiche di segnalazione iniziate dal TCR non sono da esso trasdotte: questo compito viene svolto. dalle proteine invarianti CD3 e , che insieme al recettore formano il cosiddetto complesso TCR. La. segnalazione è dunque legata a elementi estremamente variabili (il TCR) e a elementi costanti. Le cellule T esprimono altri recettori di membrana che non riconoscono l’antigene ma che contribuiscono. alla risposta: in generale si parla di molecole accessorie. Il ruolo fisiologico è per alcune. di facilitare la segnalazione del complesso TCR, per altre di fornire secondi segnali che attivano. completamente le cellule. Altre molecole accessorie ancora servono a stabilizzare il legame con le. APC, in modo da garantire il tempo necessario alla trasduzione del segnale TCR per antigeni MHC-associati. Il recettore antigenico sia dei linfociti CD4+ che dei CD8+ è un eterodimero costituito da una catena. e una tra loro unite da un ponte disolfuro. Entrambe le catene sono costituite da un dominio. simil-Ig N terminale variabile, uno simil Ig costante, un dominio transmembrana e una piccola. regione citoplasmatica; la porzione extracellulare è dunque simile alla porzione legante l’antigene di un. anticorpo, con una regione variabile e una costante sulla catena leggera e una variabile e una costante. sulla catena pesante. Le regioni variabili V delle catene del TCR contengono piccole frazioni sulle quali la grande variabilità. è concentrata: si parla di CDR, o Complementarity Determing Regions. Tre CDR sulla catena sono. giustapposte a tre regioni simili sulla catena a formare la parte del recettore che riconosce i complessi. peptide-MHC. La regione variabile della catena contiene poi una quarta regione ipervariabile che è. il sito di legame per i cosiddetti superantigeni. Ogni catena del TCR è codificata da più segmenti genici. che subiscono riarrangiamento somatico durante la maturazione dei linfociti. In entrambe le catene. la terza regione ipervariabile è composta da sequenze codificate dai segmenti genici V e J (catena ) o. dai segmenti V, D, J (catena ). Le regioni costanti C di entrambe le catene formano cerniere che contengono i residui di cisteina per. i ponti disolfuro. I TCR e gli anticorpi sono strutturalmente simili, ma vi sono delle profonde differenze. I TCR non. esistono in froma secreta e non hanno funzioni effettrici da soli, inoltre non subiscono variazioni. nella regione C e non hanno maturazione della loro affinità Ruolo del TCR nel riconoscimento dell’antigene. Il riconoscimento è mediato dalle regioni determinanti la complementarietà, o CDR, formate da entrambe. le catene del TCR. Queste catene formano un singolo recettore eterodimerico responsabile sia. della specificità per il peptide che per l’MHC. Il sito di legame per l’antigene è formato dalle sei CDR sulle due catene che formano un’interfaccia. molto simile a quella degli anticorpi. Il contatto tra il TCR e il complesso peptide-MHC è limitato a uno. o due aminoacidi: le cellule T riconoscono dunque i loro substrati sulla base di differenze minime. L’affinità del TCR per i complessi è bassa, molto più di quella della maggior parte degli anticorpi. Questa bassa affinità è probabilmente la ragione per cui sono necessarie le molecole di adesione per. avere una risposta biologica. Il TCR e le sue molecole accessorie sul linfocita T si muovono in maniera. coordinata ai loro ligandi sulle membrane delle APC per creare una struttura sopramolecolare detta. sinapsi immunologica. 1.2 Proteine CD3 e del complesso TCR. Le proteine CD3 e trasducono il segnale che porta all’attivazione del linfocita dietro stimolo del TCR. La molecola CD3 è in realtà un insieme di tre proteine designate , e . Le tre proteine CD3 e la. sono sempre uguali in tutte le cellule T: non hanno infatti ruolo nel riconoscimento ma solo nella. trasduzione del segnale Struttura. Le tre proteine CD3 sono omologhe tra loro, e le regioni extracellulari di tutte contengono un singolo. dominio simil-Ig: queste tre proteine sono dunque membri della superfamiglia delle Ig. I domini. citoplasmatici variano da 44 a 81 aminoacidi di lunghezza e ciascuno contiene una copia di una sequenza. conservata detta ITAM (Immunoreceptor Tyrosine-based Activation Motif) che gioca un ruolo. fondamentale nella segnalazione da parte del complesso. La catena ha una piccola regione extracellulare, una transmembrana e una lunga regione citoplasmatica. contentente tre ITAM. L’espressione del complesso TCR richiede la sintesi di tutti i suoi componenti. Nei linfociti T maturi. infatti l’intero complesso viene prodotto nel RE e trasportato sulla membrana Funzione. Il primo evento intracellulare a seguito del riconoscimento antigenico è la fosforilazione dei residui di. tirosina contenuti nei domini ITAM di CD3 e da parte di kinasi quali Lck o Fyn. Lck si associa alle. code citoplasmatiche di CD4 e CD8, Fyn a CD3. Le fosfotirosine così create diventano siti di attacco per. una tirosin kinasi, ZAP-70, che viene reclutata dalla catena e porterà alla variazione dell’espressione. genica delle cellule T. 1.3 Recettori antigenici dei linfociti. Il TCR è un secondo tipo di eterodimero espresso in un piccolo set di linfociti T -negativi; questo. recettore è comunque associato alle proteine CD3 e . La maggioranza delle cellule con questo recettore. non esprimono CD4 o CD8. Le cellule che esprimono questo recettore sono linearmente distinte da quelle che esprimono il più. comune TCR: in totale meno del 5% dei linfociti esprime questa struttura. Queste cellule, insieme. alle cellule NK-T, i linfociti B di tipo B-1 e le cellule MZ B, potrebbero rappresentare un importante. collegamento tra l’immunità innata e quella adattativa. Le cellule con recettore non riconoscono. gli antigeni peptidici MHC-associati e non sono MHC ristrette; alcune riconoscono piccole molecole. fosforilate o lipidi comuni nei batteri, altre riconoscono proteine che non richiedono processamento o. collaborazione delle APC. Il ruolo di queste cellule è comunque poco definito in quanto topi deficitari. non si mostrano particolarmente immunodeficienti o maggiormente suscettibili alle infezioni batteriche. 1.4 Recettori antigenici delle cellule NK-T. Una piccola popolazione di linfociti T esprime i markers tipici delle cellule NK: si tratta delle cellule. NK-T. Le catene alfa del TCR di queste cellule hanno una diversità limitata e sono caratterizzate da un. riarrangiamento caratteristico nell’uomo; queste cellule fanno ancora una volta da ponte tra l’immunità. innata e quella adattativa. Tutte i TCR delle cellule NK-T riconoscono lipidi legati alle molecole simil-MHCI CD1. Queste cellule. producono rapidamente citochine quali IL-4 e IFN- a seguito della stimolazione. 1.5 Corecettori e recettori costimolanti nelle cellule T. I corecettori sono una categoria di proteine di membrana che amplificano il segnale di TCR; queste. strutture legano le molecole di MHC. I costimolanti conducono anch’essi dei segnali che attivano le. cellule T, ma riconoscono molecole sulle APC che non sono parte del complesso MHC-peptide CD4 e CD8: Corecettori coinvolti nell’attivazione delle cellule T MHC- ristrette. Le cellule T mature esprimono CD4 o CD8, ma mai entrambi. Queste strutture interagiscono con. entrambe le classi di MHC quando i TCR della cellula riconoscono i complessi MHC-peptide dell’APC. La funzione principale è nella trasduzione del segnale al momento del riconoscimento, ma possono. 3. anche aumentare l’efficacia del legame tra cellula T ed APC. Nel pool dei linfociti maturi circa il 65% esprime CD4 e il 35% CD8. Struttura Entrambi i corecettori sono glicoproteine transmembrana facenti parte della superfamiglia. Ig. CD4 viene espresso come monomero e presenta quattro domini simil-Ig extracellulari, una regione. transmembrana e una coda basica citoplasmatica. I due domini simil-Ig amino terminali del CD4. legano il dominio 2 non polimorfico dell’MHCII. Le molecole CD8 esistono quasi sempre sotto forma di eterodimeri di due catene dette CD8 e. CD8. Entrambe presentano un singolo dominio Ig extracellulare, una regione transmembrana e la. coda citoplasmatica basica. Il dominio Ig di CD8 lega il dominio 2 non polimorfico delle molecole di. MHCI. Funzione La separazione delle risposte dei linfociti CD4+ e CD8+ è dovuta alla capacità di queste. molecole di legare solamente una classe di MHC e non l’altra. CD4 è in grado di legare MHCII e viene espresso sui linfociti i cui TCR riconoscono i complessi. peptidici di questo tipo. Quasi tutti i linfociti CD4+ sono cellule di supporto che producono. citochine. CD8 è in grado di legare le molecole MHCI. Quasi tutti i linfociti CD8+ sono linfociti citotossici il. cui ruolo è sradicare le infezioni intracellulari. Esistono linfociti CD4+ con funzioni citotossiche. ma sono comunque MHCII ristretti. CD4 e CD8 partecipano ai primi eventi segnalatori dopo il riconoscimento del complesso MHC-peptide. Queste funzioni sono mediate da una tirosin chinasi specifica dei linfociti T che prende il nome di. Lck: questo enzima è associato in modo non covalente alle code sia del CD4 che del CD8. Quando un. linfocita riconosce il complesso MHC l’interazione di CD4/8 con l’MHC porta il corecettore e la sua Lck. nelle vicinanze del CTR; Lck a questo punto fosforila i domini ITAM delle proteine CD3 e e da il via. alla segnalazione Recettori costimolanti ed inibitori della famiglia CD28. CD28 è una proteina che trasduce il segnale in associazione ai segnali in arrivo dal complesso TCR per. attivare le cellule T naive. I linfociti T naive in generale necessitano di due segnali extracellulari. distinti per proliferare e differenziare; il primo deriva dal legame dell’antigene al recettore e garantisce. la specificità della risposta. Il secondo segnale viene fornito da molecole che vengono definite in. generale costimolatori. I costimolatori meglio definiti per i linfociti T sono chiamati B7-1 (CD80) e B7-2 (CD86) e sono. espressi sulle cellule dendritiche, sui macrofagi e sui linfociti B; queste molecole hanno specifici recettori. sul linfocita. Il primo recettore per B7 scoperto fu la molecola CD28, espressa su quasi tutti i. CD4+ e su metà dei CD8+. Il legarsi delle molecole B7 delle APC a CD28 fornisce al linfocita il segnale. per esprimere proteine anti apoptosi, per produrre fattori di crescita e citochine e per promuovere. proliferazione e differenziazione. 4. Un secondo recettore per molecole B7 venne successivamente scoperto e chiamato CTLA-4; questa. struttura è omologa a CD28 e viene espressa sui linfociti recentemente attivati: la sua funzione è inibire. l’attivazione controbilanciando i segnali in arrivo dal complesso TCR e da CD28. Molte altre strutture. sono state scoperte in grado di legare le molecole B7 e sono equamente divise tra vie di attivazione e vie. di terminazione CD2 e la famiglia SLAM di recettori costimolanti. Un’importante famiglia di proteine che gioca un ruolo nell’attivazione delle cellule T ed NK è un gruppo. di proteine strutturalmente legate ad un recettore detto CD2. Questo recettore contiene due domini. Ig extracellulari, una regione di membrana e una lunga coda citoplasmatica. Nell’uomo il principale. ligando è la molecola LFA-3 (Leukocyte Function-associated Antigen 3) che è espressa in molte cellule. ematopoietiche e non. CD2 è esempio di molecola accessoria che funziona sia come una molecola di. adesione che come trasduttore del segnale. Un sottogruppo distinto di protine CD2 è detto SLAM (Signaling Lymphocytic Activation Molecule). Le SLAM sono proteine integrali di membrana con due domini Ig extracellulari e una coda citoplasmatica. che contiene un dominio detto ITSM (Immunoreceptor Tyrosin-based Switch Motif); il dominio. si lega ad un adattatore detto SAP (SLAM Associated Protein) che contiene un dominio SH2 in grado. di fare da ponte tra SLAM e Fyn, una chinasi. Un importante membro della famiglia SLAM è 2B4, mutazioni in questa molecola possono seriamente danneggiare il sistema immunitario Altre molecole accessorie dei linfociti T. CD44 è una glicoproteina espressa in varie cellule, tra cui linfociti T maturi, timociti, cellule B, granulociti, macrofagi, eritrociti e fibroblasti. Questa molecola lega lo ialuronato e questa proprietà è. responsabile della detenzione dei linfociti T nei siti extravascolari di infezione. Le cellule CD4+ attivate esprimono una proteina della famiglia del TNF detta CD40L, che lega CD40. dei linfociti B e di altre cellule e le attiva. CD40L è quindi un importante mediatore delle funzioni helper. di questi linfociti. I linfociti attivati esprimono anche il FAS ligand; l’attivazione di FAS da parte di FASL porta. all’apoptosi ed è importante per eliminare le cellule T iperstimolate. 5.")
9
Il TCR è una molecola espressa sui linf
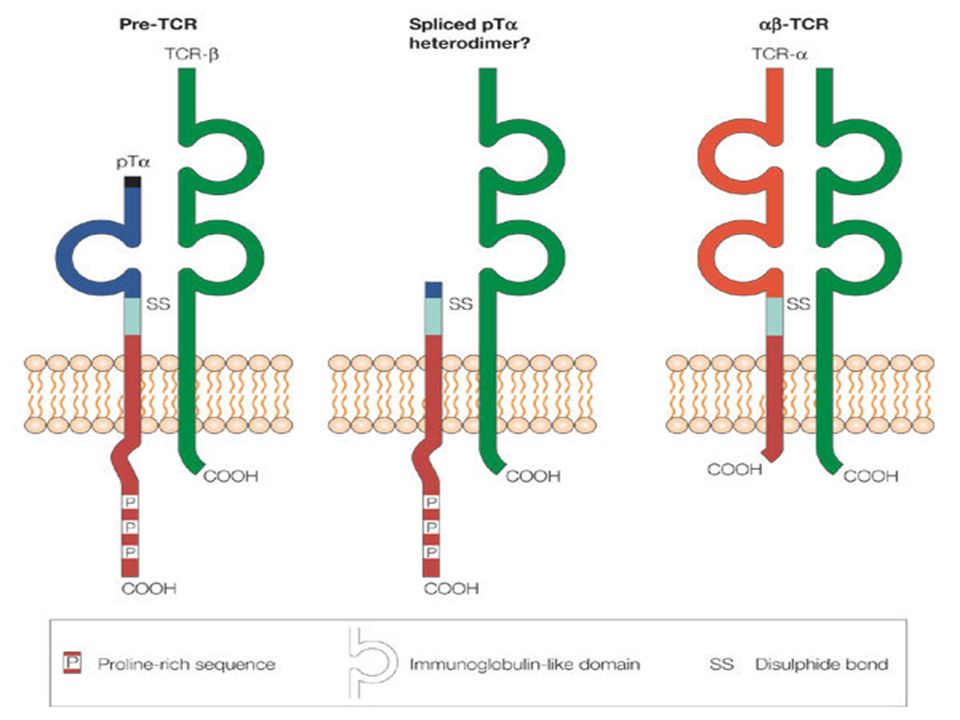
Il TCR è una molecola espressa sui linf. T riconosce Ag espressi sull’MHC . E’ un eterodimero, composto da 2 sub-unità α/β, e da sub-unità δ/γ. Ogni linf. T durante il suo sviluppo va incontro a riarrangiamento genico, che gli permette di avere un TCR con una specificità singola verso un unico Ag, su una gamma di possibilità di Lo studio della sub-unità β serve a valutare se in un pool di cellule T sono presenti espansioni clonali. La regione V-β è parte di questa ricombinazione. Le possibili famiglie della regione V-β umana sono 24, ma le V-β-10 e V-β-19 non sono funzionali. Valutando se l’espressione delle V-β è policlonale, oligoclonale o monoclonale si possono avere informazioni sulla modalità con cui il comparto linfocitario T sta rispondendo ad uno stimolo antigenico. Infatti, il linfocita T che riconosce l’Ag si attiva e comincia a replicarsi ( cloni ).
.")
11
Segnale per iniziare la risposta immune
Ag internalizzato e complessato con MHC Complesso peptide-MHC interagisce con TCR

13
Il complesso maggiore di istocompatibilità di classe I- HLA di classe I
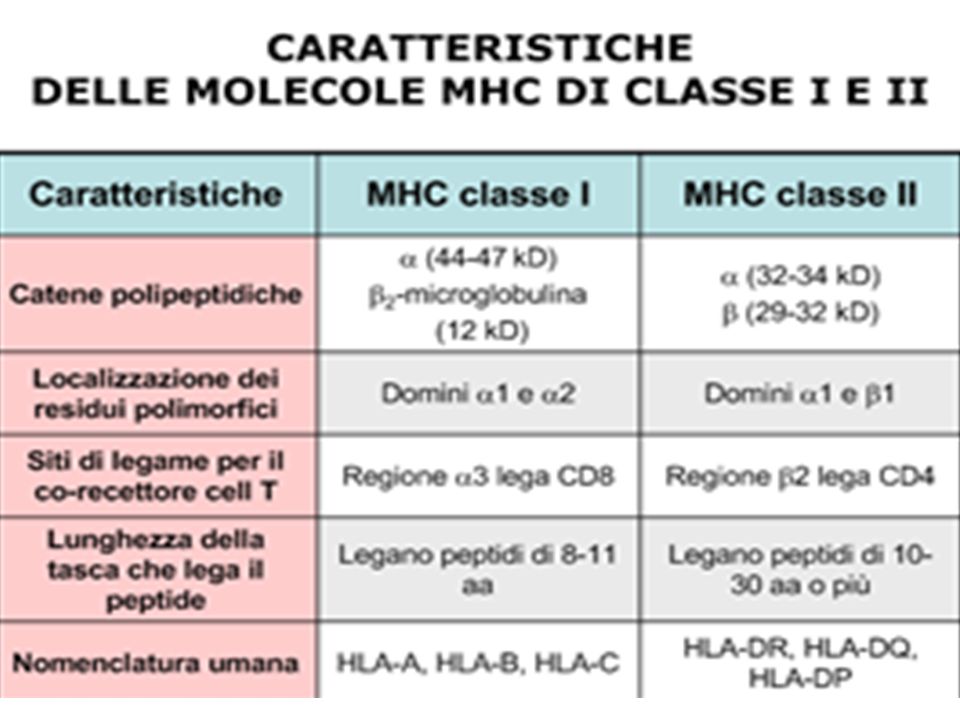
E’ formato da 4 catene pepdidiche: 3 catene α sono legate ad 1 catena β non a contatto con la membrana cellulare. Le catene α-1 e α-2, lunghe circa 90 aa, formano una tasca, dove viene esposto l‘Ag non-self derivato da proteine virali o tumorali frammentate. Tale zona è responsabile del riconoscimento, da parte dei linfociti T, delle cellule sane dalle cellula infette o tumorali. Alle estremità della catena ci sono 2 ac. carbossilici COOH. La catena β2, non va all'interno del citoplasma e non è adiacente alla membrana. Il complesso HLA di classe I, deriva da tre loci sul cromosoma 6 chiamati HLA-A, HLA-B, HLA-C. La catena β-2, invece, è codificata nel cromosoma 15. Il complesso maggiore di istocompatibilità di classe I, che nell'uomo prende il nome di HLA di classe I, ha una struttura particolare in quanto risulta formato da quattro catene di natura pepdidica. Tre catene alpha sono legati ad una catena beta che non ha contatto con la membrana cellulare. Le catene alpha-1 e alpha-2, della lunghezza di circa 90 amminoacidi, formano una fessura che viene definita tasca. Nella zona della tasca viene esposto l'antigene non-self derivato dalla frammentazione endocellulare di proteine virali o tumorali ed è quindi tale zona responsabile del riconoscimento, da parte dei linfociti T, delle cellule sane dalle cellula infette o tumorali. Le estremità della catena sono definite da due acidi carbossilici COOH. La catena beta-2, pur avendo una simile struttura proteica, non sembra penetrare all'interno del citoplasma e non è neppure adiacente alla membrana. Il complesso LHA di classe I è stato il primo ad essere scoperto e, geneticamente, deriva da tre loci situati sul cromosoma 6 chiamati HLA-A, HLA-B, HLA-C. La catena beta-2, a differenza delle altre tre, non è codificata nel cromosoma 6 ma nel cromosoma 15. Genetica dell'LHA di classe I. L'informazione genetica necessaria per codificare l'HLA di classe I è presente nel cromosoma 6 e, più precisamente, nella zona che va verso il telomero a differenza di quanto avviene nell'HLA di classe II dove i loci sono localizzati nella partecentromerica. Nella zona si distinguono tre loci definiti HLA-A, HLA-B e HLA-C.

15
Il complesso maggiore di istocompatibilità di classe I- HLA di classe I
L'HLA di classe II è presente solo nei linf. B, nelle cellule T attivate, e nelle cellule presentanti l'antigene (APC). Struttura dell' HLA di classe II. La struttura dell'HLA di classe II è simile alla classe I. La tasca è localizzata nell'insenatura che si forma tra le catene α-1 e β-1. Le estremita dell'HLA attraversano la membrana citoplasmatica per inserirsi all'interno del citosol, a differenza di quanto avviene nell'HLA di classe I dove soltanto la catena α-3 si insedia all'interno del citoplasma. Ambedue estremità della catena possiedono due terminazioni carbossilche. Genetica dell'HLA di classe II. E’ codificato nel cromosoma 6. Si definiscono tre regioni altamente variabili denominate DPA, DQA e DRA per le catene α e DPB, DQB e DRB per le catene β.
. Struttura dell HLA di classe II. La struttura dell HLA di classe II è simile alla classe I. La tasca è localizzata nell insenatura che si forma tra le catene α-1 e β-1. Le estremita dell HLA attraversano la membrana citoplasmatica per inserirsi all interno del citosol, a differenza di quanto avviene nell HLA di classe I dove soltanto la catena α-3 si insedia all interno del citoplasma. Ambedue estremità della catena possiedono due terminazioni carbossilche. Genetica dell HLA di classe II. E’ codificato nel cromosoma 6. Si definiscono tre regioni altamente variabili denominate DPA, DQA e DRA per le catene α e DPB, DQB e DRB per le catene β.")
18
I linf.T sono divisi in 2 grandi categorie distinguibili per le diverse funzioni e le proteine di membrana CD4 e CD8 (co-recettori) Durante il riconoscimento dell’Ag, CD4 o CD8 si associano al recettore di membrana e legano i siti costanti della porzione MHC del complesso peptide: MHC CD4 è una molecola a catena singola composta da 4 domini simili a quelli delle Ig. I primi due formano una bacchetta rigida unita a una cerniera flessibile l’altra bacchetta formata da D3 e D4 lega L’MHC II

19
La capacità del sistema immunitario di differenziare il self dal non-self è in larga parte determinata dai prodotti del complesso maggiore di istocompatibilità (Major Histocompatibility Complex, MHC), i cui geni si trovano sul cromosoma 6, appartengono alla superfamiglia dei geni delle Ig e sono soggetti a ricombinazione genica. I prodotti del MHC di classe I sono costituiti dagli HLA-A, -B e-C; essi sono ampiamente distribuiti nell'organismo e sono presenti sulla superficie di tutte le cellule nucleate e sulle piastrine. I prodotti del MHC di Classe II sono costituiti dagli HLA-D, -DR, -DP e-DQ; essi hanno una distribuzione più limitata sulle cellule B, sui macrofagi, sulle cellule dendritiche, sulle cellule di Langerhans e sulle cellule T attivate (ma non su quelle quiescenti). Le cellule B sono in grado di rispondere agli Ag solubili, ma le cellule T lo fanno raramente e riconoscono l'Ag solamente quando è associato al MHC; esse riconoscono quindi il complesso MHC/Ag. Il meccanismo attraverso il quale l'Ag viene processato e associato al MHC prima di essere presentato alle cellule T viene realizzato dalle cellule di presentazione dell'antigene (Antigen-Presenting Cells, APC), p. es. le cellule di Langerhans, i monociti, i macrofagi, le cellule dendritiche follicolari e le cellule B. Sebbene i particolari non siano pienamente compresi, sembra che per essere processato l'Ag debba essere esposto, degradato e frammentato. Nel caso della presentazione esogena, l'Ag viene sottoposto a endocitosi e degradazione all'interno dei lisosomi, viene associato ai prodotti del MHC di classe II e viene trasportato fino alla superficie cellulare. Nel caso della presentazione endogena, l'Ag viene prodotto intracellularmente (p. es. da un'infezione virale) e viene sottoposto a degradazione al di fuori dei lisosomi, all'interno di organuli chiamati proteosomi. I peptidi che ne risultano vengono trasferiti al reticolo endoplasmatico rugoso (RER) per mezzo di proteine di trasporto. Una volta all'interno del RER, questi peptidi vengono associati con i prodotti del MHC di classe I per poi essere trasportati fino alla superficie cellulare. È importante sapere se l'Ag viene associato con il MHC di classe I o di classe II, perché le molecole CD4 e CD8 agiscono come molecole di adesione accessorie legandosi rispettivamente agli Ag di classe II o di classe I. L'interazione del TCR con il complesso MHC/Ag può non essere sufficiente per indurre l'attivazione delle cellule T. È necessaria la presenza di un segnale di coattivazione; questo secondo segnale è mediato dall'interazione del CD28 presente sulla superficie delle cellule T con il CD80 o il CD86 presente sulle APC. L'assenza dell'interazione CD28/CD80-CD86 può rendere la cellula T anergica o tollerante (v. Fig. 146-1).
, p. es. le cellule di Langerhans, i monociti, i macrofagi, le cellule dendritiche follicolari e le cellule B. Sebbene i particolari non siano pienamente compresi, sembra che per essere processato l Ag debba essere esposto, degradato e frammentato. Nel caso della presentazione esogena, l Ag viene sottoposto a endocitosi e degradazione all interno dei lisosomi, viene associato ai prodotti del MHC di classe II e viene trasportato fino alla superficie cellulare. Nel caso della presentazione endogena, l Ag viene prodotto intracellularmente (p. es. da un infezione virale) e viene sottoposto a degradazione al di fuori dei lisosomi, all interno di organuli chiamati proteosomi. I peptidi che ne risultano vengono trasferiti al reticolo endoplasmatico rugoso (RER) per mezzo di proteine di trasporto. Una volta all interno del RER, questi peptidi vengono associati con i prodotti del MHC di classe I per poi essere trasportati fino alla superficie cellulare. È importante sapere se l Ag viene associato con il MHC di classe I o di classe II, perché le molecole CD4 e CD8 agiscono come molecole di adesione accessorie legandosi rispettivamente agli Ag di classe II o di classe I. L interazione del TCR con il complesso MHC/Ag può non essere sufficiente per indurre l attivazione delle cellule T. È necessaria la presenza di un segnale di coattivazione; questo secondo segnale è mediato dall interazione del CD28 presente sulla superficie delle cellule T con il CD80 o il CD86 presente sulle APC. L assenza dell interazione CD28/CD80-CD86 può rendere la cellula T anergica o tollerante (v. Fig ).")
22
Le cellule infettate da microbi intracellulari, es virus, vengono catturate dalle APCs, in particolare da cellule dentritiche, fagocitate all’interno del fagosoma gli Ag di virus vengono tagliati e presentati sulla superficie in associazione con le molecole di MHC. I linf T riconoscono gli antigeni microbici e le molecolae costimolatrici presenti sulle APCs e quindi vengono attivato Le cellule infettate da microbi intracellulari, es. virus, vengono catturate dalle APCs, in particolare da cellule dentritiche, fagocitate all’interno del fagosoma gli Ag di virus vengono tagliati e presentati sulla superficie in associazione con le molecole di MHC. I linf T riconoscono gli antigeni microbici e le molecole co-stimolatrici presenti sulle APCs e quindi vengono attivati

23
Via dell’MHC I: gli Ag proteici all’interno del citosol vengono processati dai proteosomi e trasportati nel RE dove incontrano e si legano alle molecole di MHC classe I

24
Via delle Molecole MHC II: Ag proteici extracellulari vengono catturati per endocitosi nelle vescicole. All’interno vengono processati e i peptidi si legano alle molecole di MHCII: Via dell’MHC I gli Ag proteici all’interno del citosol vengono processati dai proteosomi e trasportati nel RE dove incontrano e si legano alle molecole di MHCI Via delle Molecole MHC II: Ag proteici extracellulari vengono catturati per endocitosi nelle vescicole. All’interno vengono processati e i peptidi si legano alle molecole di MHCII

27
Entrambi i corecettori sono glicoproteine transmembrana
appartenenti alla superfamiglia Ig. CD4 viene espresso come monomero presenta quattro domini simil-Ig extracellulari, una regione transmembrana e una coda basica citoplasmatica. I due domini simil Ig amino terminali del CD4 legano il dominio 2 non polimorfico dell’MHCII. Le molecole CD8 esistono quasi sempre sotto forma di eterodimeri di due catene dette CD8 e CD8. Entrambe presentano un singolo dominio Ig extracellulare, una regione transmembrana e la coda citoplasmatica basica. Il dominio Ig di CD8 lega il dominio 2 non polimorfico delle molecole di MHCI. Struttura Entrambi i corecettori sono glicoproteine transmembrana facenti parte della superfamiglia Ig. CD4 viene espresso come monomero e presenta quattro domini simil-Ig extracellulari, una regione transmembrana e una coda basica citoplasmatica. I due domini simil-Ig amino terminali del CD4 legano il dominio 2 non polimorfico dell’MHCII. Le molecole CD8 esistono quasi sempre sotto forma di eterodimeri di due catene dette CD8 e CD8. Entrambe presentano un singolo dominio Ig extracellulare, una regione transmembrana e la coda citoplasmatica basica. Il dominio Ig di CD8 lega il dominio 2 non polimorfico delle molecole di MHCI.

28
Signal transduction through the T cell receptor The T cell receptor (TCR) is a complex of integral membrane proteins that participates in the activation of T cells in response to the presentation of antigen. Specific recognition and binding by the clonotype-specific a/b heterodimer leads to activation of transcription and commitment of the T cell to CD4+ or CD8+ fate. This activation involves other subunits of the receptor complex as well as other membrane-associated molecules that couple the extracellular liganding event to downstream signaling pathways such as protein phosphorylation, the release of inositol phosphates and the elevation of intracellular calcium levels. This discussion of TCR signaling focusses on the involvement of TM domains, rather than on providing a complete overview. The intracellular portions of the CD3 g, d, e, and z subunits contain copies of a sequence motif termed ITAMs (immunoreceptor tyrosine-based activation motifs). ITAMs can serve as protein tyrosine kinase substrates and, after phosphorylation, as binding sites for SH2 domains of yet other kinases. The regulation and mechanism of the recruitment of protein kinases to the activated T cell receptor is being studied intensively; members of both the Syk family (ZAP-70) and Src family (Lck) of kinases are involved in this process. A model outlining our current knowledge of TCR and linker for activation of T cells (LAT) signalling complexes is illustrated. Following TCR ligation, SRC-family protein tyrosine kinases (PTKs) — for example, LCK and FYN — are activated, resulting in phosphorylation of CD3 modules of the TCR complex and activation of SYK-family PTKs —for example, -chain-associated protein of 70kD (ZAP70). Activated ZAP70 phosphorylates LAT and SLP76 (SRC-homology 2 (SH2)-domain-containing leukocyte protein of 76 kD). Tyrosine-phosphorylated LAT then recruits several SH2-domain-containing proteins, including growth factor receptor-bound protein 2 (GRB2), GRB2-related adaptor protein (GADS) and phospholipase C-1 (PLC-1). Through its constitutive association with GADS, SLP76 is also recruited to LAT following TCR stimulation. Evidence indicates that SLP76 also constitutively associates with the SH3 domain of PLC-1. Activation of PLC-1 results in the hydrolysis of phosphatidylinositol 4,5-bisphosphate to inositol 3,4,5-triphosphate (IP3) and diacylglycerol (DAG). IP3 production leads to increases of intracellular free Ca2+ concentration, whereas DAG can activate both protein kinase C- (PKC-) and RAS guanyl nucleotide-releasing protein (RASGRP). Phosphorylated LAT also recruits the SH2 domain of GRB2, and therefore, the GRB2-associated RAS guanosine nucleotide-exchange factor (GEF), son-of-sevenless (SOS), thereby providing an additional possible mechanism of RAS activation through LAT. Tyrosine-phosphorylated SLP76 also associates with the RHO-family GEF, VAV1, and the adaptor protein, NCK. A trimolecular complex between SLP76, VAV1 and NCK-associated p21-activated kinase 1 (PAK1) has been proposed as a potential mechanism for SLP76 regulation of actin cytoskeletal rearrangements following TCR stimulation. APC, antigen-presenting cell; IL-2, interleukin-2; MAPK, mitogen-activated protein kinase
 signalling complexes is illustrated. Following TCR ligation, SRC-family protein tyrosine kinases (PTKs) — for example, LCK and FYN — are activated, resulting in phosphorylation of CD3 modules of the TCR complex and activation of SYK-family PTKs —for example, -chain-associated protein of 70kD (ZAP70). Activated ZAP70 phosphorylates LAT and SLP76 (SRC-homology 2 (SH2)-domain-containing leukocyte protein of 76 kD). Tyrosine-phosphorylated LAT then recruits several SH2-domain-containing proteins, including growth factor receptor-bound protein 2 (GRB2), GRB2-related adaptor protein (GADS) and phospholipase C-1 (PLC-1). Through its constitutive association with GADS, SLP76 is also recruited to LAT following TCR stimulation. Evidence indicates that SLP76 also constitutively associates with the SH3 domain of PLC-1. Activation of PLC-1 results in the hydrolysis of phosphatidylinositol 4,5-bisphosphate to inositol 3,4,5-triphosphate (IP3) and diacylglycerol (DAG). IP3 production leads to increases of intracellular free Ca2+ concentration, whereas DAG can activate both protein kinase C- (PKC-) and RAS guanyl nucleotide-releasing protein (RASGRP). Phosphorylated LAT also recruits the SH2 domain of GRB2, and therefore, the GRB2-associated RAS guanosine nucleotide-exchange factor (GEF), son-of-sevenless (SOS), thereby providing an additional possible mechanism of RAS activation through LAT. Tyrosine-phosphorylated SLP76 also associates with the RHO-family GEF, VAV1, and the adaptor protein, NCK. A trimolecular complex between SLP76, VAV1 and NCK-associated p21-activated kinase 1 (PAK1) has been proposed as a potential mechanism for SLP76 regulation of actin cytoskeletal rearrangements following TCR stimulation. APC, antigen-presenting cell; IL-2, interleukin-2; MAPK, mitogen-activated protein kinase.")
Presentazioni simili












 SPECIFICA/SELETTIVA:>")



>")










