Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Esame di Tecniche Biomolecolari
Modulo: Clonaggio ed Espressione Docente: Dott. Pizzo Ricevimento: Martedì ore 10: :00 Contatti:

2
Programma Obiettivi: approfondimento delle conoscenze teorico-metodologiche necessarie per progettare e condurre produzioni di proteine ricombinanti in diversi ospiti Contenuti: Clonaggio genico. Espressione di proteine ricombinanti in procarioti. Vettori di espressione procariotici di ultima generazione. Fattori che influenzano l’efficienza di espressione in ospiti batterici: caratteristiche del ceppo selezionato, temperatura, mezzo di coltura, induttori, codon usage. -Espressione di proteine ricombinanti in eucarioti. Vettori di espressione eucariotici. Espressione in lievito. Espressione in piante. Analisi in silico di sequenze proteiche per la scelta del sistema di espressione ottimale: software per predizioni di solubilità, stabilità, sensibilità alle proteasi, presenza di domini transmembrana, destino cellulare. Diversi sistemi di produzione di proteine ricombinanti a confronto: vantaggi e svantaggi dell’utilizzo di batteri, lieviti, cellule vegetali e cellule di mammifero

3
ISCRIZIONE AI LABORATORI 2° SEMESTRE A.A. 2013/2014
Accedere al seguente sito webdocenti: Fornire: USERNAME: matricola N82/numero a 6 cifre PASSWORD: PIN numerico a 5 cifre Nella sezione Iscrizione ai gruppi/test prenotarsi cliccando sull’icona “effettua iscrizione” o dalla prima pagina su “clicca qui per iscriverti” E’ importante confermare l’iscrizione cliccando su conferma e attendere che essa sia avvenuta con successo. Nella sezione Avvisi si potranno apprendere varie informazioni di carattere generale. Nella sezione Orario delle Lezioni è visualizzabile la data e l’orario di ciascuna esercitazione (come da tabella dell’orario delle lezioni). Una volta scadute le iscrizioni, la suddivisione nei vari sottogruppi sarà pubblicata nella sezione Gestione materiale didattico.
. Una volta scadute le iscrizioni, la suddivisione nei vari sottogruppi sarà pubblicata nella sezione Gestione materiale didattico.")
4
Cloning - a definition From the Greek - klon, a twig
An aggregate of the asexually produced progeny of an individual;a group of replicas of all or part of a macromolecule (such as DNA or an antibody) An individual grown from a single somatic cell of its parent & genetically identical to it Clone: a collection of molecules or cells, all identical to an original molecule or cell
 An individual grown from a single somatic cell of its parent & genetically identical to it. Clone: a collection of molecules or cells, all identical to an original molecule or cell.")
5
What is DNA cloning? When DNA is extracted from an organism, all its genes are obtained In gene (DNA) cloning a particular gene is copied (cloned)
 cloning a particular gene is copied (cloned) .")
6
What is Gene cloning? To "clone a gene" is to make many copies of it
Act of making many identical copies of gene Gene can be an exact copy of a natural gene Gene can be an altered version of a natural gene

7
Il clonaggio Clonaggio: viene generata una molecola di DNA ricombinante, cioe’ isolata dal suo contesto e introdotta in un replicone (una unita’ di DNA capace di replicarsi detta vettore). Questa molecola di DNA ricombinante viene introdotta in un sistema cellulare appropriato. Tutti i discendenti di questa singola cellula, detta clone, conterranno la medesima molecola di DNA ricombinante originariamente isolata, permettendo di produrne quantita’ praticamente illimitate
. Questa molecola di DNA ricombinante viene introdotta in un sistema cellulare appropriato. Tutti i discendenti di questa singola cellula, detta clone, conterranno la medesima molecola di DNA ricombinante originariamente isolata, permettendo di produrne quantita’ praticamente illimitate.")
8
Why Clone DNA? A particular gene can be isolated and its nucleotide sequence determined Control sequences of DNA can be identified & analyzed Protein/enzyme/RNA function can be investigated Mutations can be identified, e.g. gene defects related to specific diseases Organisms can be ‘engineered’ for specific purposes, e.g. insulin production, insect resistance, etc.

9
del batterio. VETTORI: PLASMIDI
Molecole extra-cromosomali di DNA circolare capaci di replicarsi e di segregare autonomamente rispetto al DNA cromosomale batterico. Presenti in natura (batteri, lieviti, alcuni eucarioti superiori) in stato di relazione parassitica o simbiontica verso la cellula ospite. Portatori di geni che arrecano un vantaggio selettivo alla cellula ospite (es. resistenza a un antibiotico) Possono ospitare DNA estraneo (con limiti di grandezza) Possono essere facilmente purificati dal DNA cromosomale del batterio.
 in stato di relazione parassitica o simbiontica verso la cellula ospite. Portatori di geni che arrecano un vantaggio selettivo alla cellula ospite (es. resistenza a un antibiotico) Possono ospitare DNA estraneo (con limiti di grandezza) Possono essere facilmente purificati dal DNA cromosomale. del batterio.")
10
Caratteristiche essenziali dei vettori plasmidici:
1) Origine di replicazione in E. Coli 2) Marcatore di selezione che permette ai batteri trasformati di crescere su terreno selettivo
 Origine di replicazione in E. Coli. 2) Marcatore di selezione che permette ai. batteri trasformati di crescere su terreno. selettivo.")
11
Solo i batteri che hanno integrato un plasmide codificante il gene per
Resistenza agli antibiotici: Solo i batteri che hanno integrato un plasmide codificante il gene per la resistenza possono crescere su agar nutritivo in presenza di antibiotico

12
Cosa sono gli enzimi di restrizione
Sono enzimi che tagliano la doppia elica del DNA in corrispondenza di specifiche sequenze. Per questo motivo vengono anche detti endonucleasi di restrizione (invece le esonucleasi distruggono il DNA partendo dalle estremità). Ne esistono molti tipi diversi, ognuno dei quali riconosce una sequenza particolare.
. Ne esistono molti tipi diversi, ognuno dei quali riconosce una sequenza particolare.")
13
Enzimi di restrizione Associazione dei frammenti per complementarietà
Taglio Frammento con estremità coesive Associazione dei frammenti per complementarietà DNA ricombinante Formazione di una molecola circolare Dna ligasi

14
Gli enzimi di restrizione proteggono i batteri dall’introduzione di molecole di DNA esogeno
Batterio vivo Batterio ceppo A Batteriofagi Batterio ceppo B Batterio lisato

15
-Nel 1970 Hamilton Smith ha isolato il primo enzima che taglia il
DNA a livello di una sequenza nucleotidica specifica. -Endonucleasi di restrizione e metiltransferasi formano il SISTEMA DI MODIFICAZIONE E RESTRIZIONE dei batteri.

16
ENDONUCLEASI DI RESTRIZIONE DI TIPO II
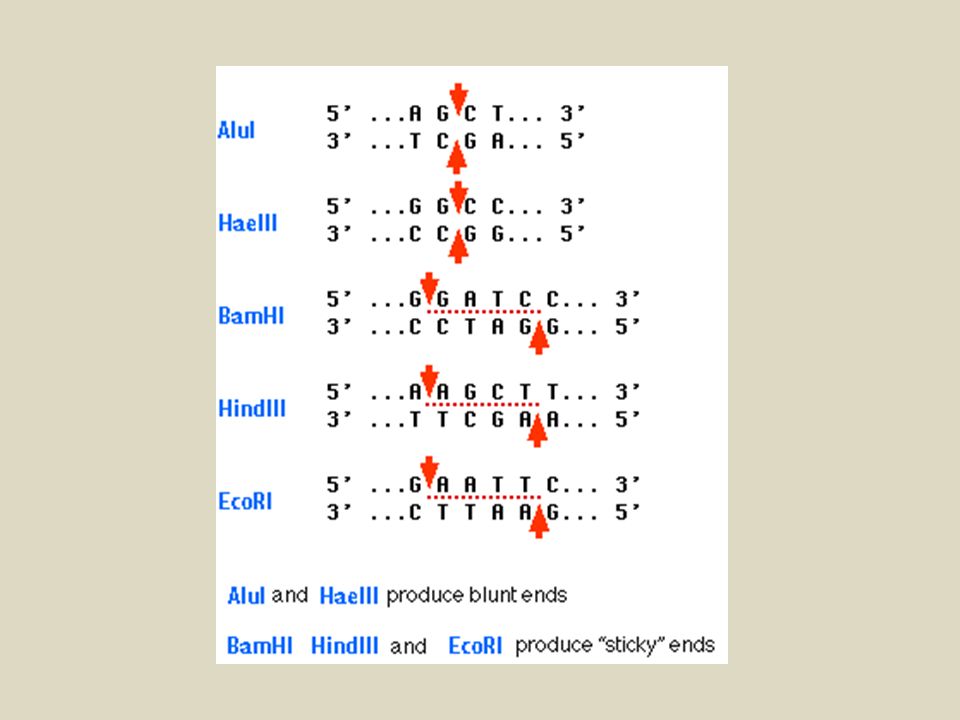
-Servono a tagliare il DNA in modo preciso e prevedibile -Sono stati isolati più di 1200 enzimi da procarioti -Riconoscono più di 130 diverse sequenze nucleotidiche (sequenze palindromiche di 4,6 o 8 nucleotidi). Hae III 5’ GGCC 3’ 3’ CCGG 5’ Eco RI 5’ GAATTC 3’ 3’ CTTAAG 5’ Not I 5’ GCGGCCGC 3’ 3’ CGCCGGCG 5’
. Hae III. 5’ GGCC 3’ 3’ CCGG 5’ Eco RI. 5’ GAATTC 3’ 3’ CTTAAG 5’ Not I. 5’ GCGGCCGC 3’ 3’ CGCCGGCG 5’")
17
Esempi di enzimi di restrizione

18
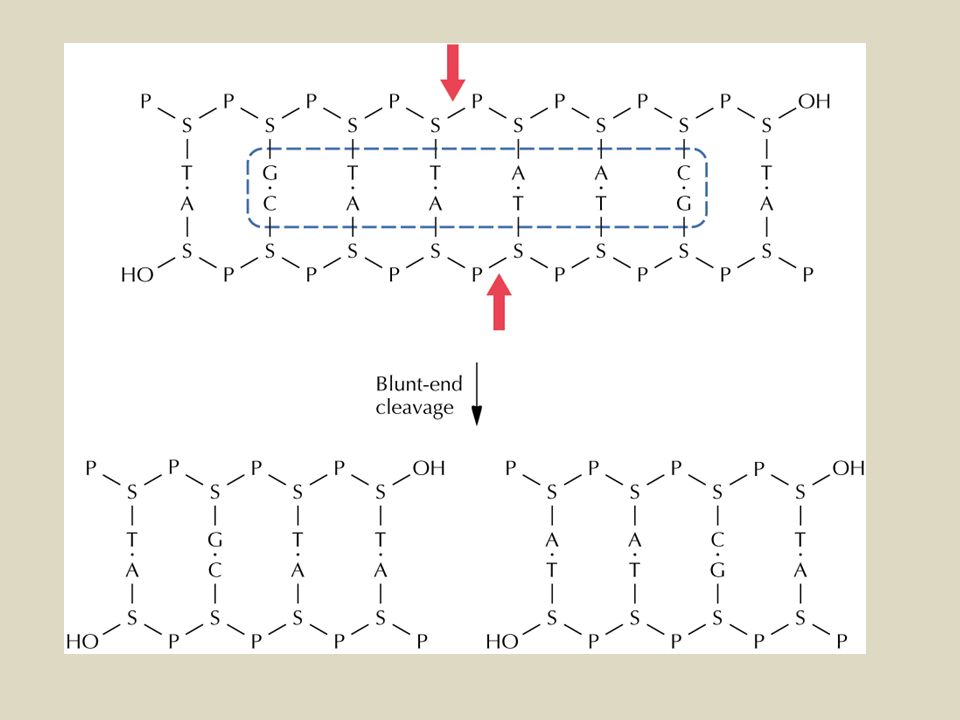
Il taglio con gli enzimi di restrizione può generare diversi tipi di estremità.
Eco RI= 5’ protruding -GAATTC G3’ 5’AATTC- -CTTAAG CTTAA5’ 3’G- Eco RV= blunt -GATATC GAT3’ 5’ATC- -CTATAG CTA5’ 3’TAG- Pst I= 3’ protruding -CTGCAG CTGCA3’ 5’G- -GACGTC G5’ 3’ACGTC-

20
Sticky end

22
DNA ligase covalently links two DNA strands
5’ 3’ Restriction enzyme Ligase

25
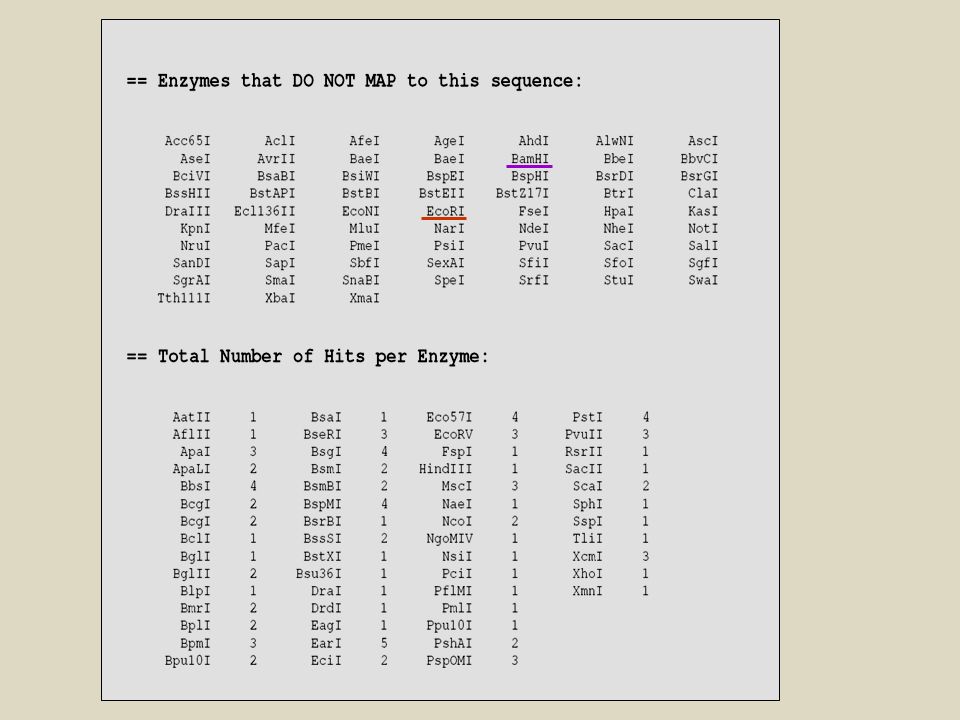
QUANTE VOLTE TAGLIA UN ENZIMA DI RESTRIZIONE?
La maggior parte riconosce palindromi di 4 o 6 nucleotidi se assumiamo che i 4 nucleotidi siano distribuiti a caso nelle molecole di DNA: -per enzimi che riconoscono palindromi di 4 nt si avrà IN MEDIA un taglio ogni 256 nucleotidi (4x4x4x4). -per enzimi che riconoscono palindromi di 6 nucleotidi avremo IN MEDIA un taglio ogni nucleotidi (46).
. -per enzimi che riconoscono palindromi di 6 nucleotidi avremo IN. MEDIA un taglio ogni nucleotidi (46).")
26
Separazione di frammenti di DNA (o RNA) di diversa lunghezza tramite elettroforesi
 di diversa lunghezza tramite elettroforesi")
27
Cella elettroforetica per gel di agarosio

28
- + Plasmide (P) M P P+B P+E G+E Eco RI (E) 10 kb Bam HI (B) Eco RI
Il DNA digerito con gli enzimi di restrizione può essere analizzato mediante elettroforesi Plasmide (P) - M P P+B P+E G+E Eco RI (E) 10 9 10 kb Bam HI (B) 8 7 6 Eco RI 5 DNA genomico (G) 3x 109 basi 4 3 2 + 1
 - M. P. P+B. P+E. G+E. Eco RI (E) kb. Bam HI (B) Eco RI. 5. DNA genomico (G) 3x 109 basi")
29
- + M MAPPATURA ENZIMATICA DI UN TRATTO DI DNA 6.6 bp PstI HindIII
1 2 3 4 + 0,5 PstI+HindIII 1.1 1.4 4.1 - 1.7 2.3 2.6 0.6 1.2 PstI HindIII 1.1 kb kb kb kb kb

30
Gli enzimi di restrizione, consentendo di tagliare il DNA in frammenti non casuali, la cui grandezza dipende unicamente dalla sequenza, costituiscono un formidabile strumento sia per l’analisi che per la manipolazione del DNA.

31
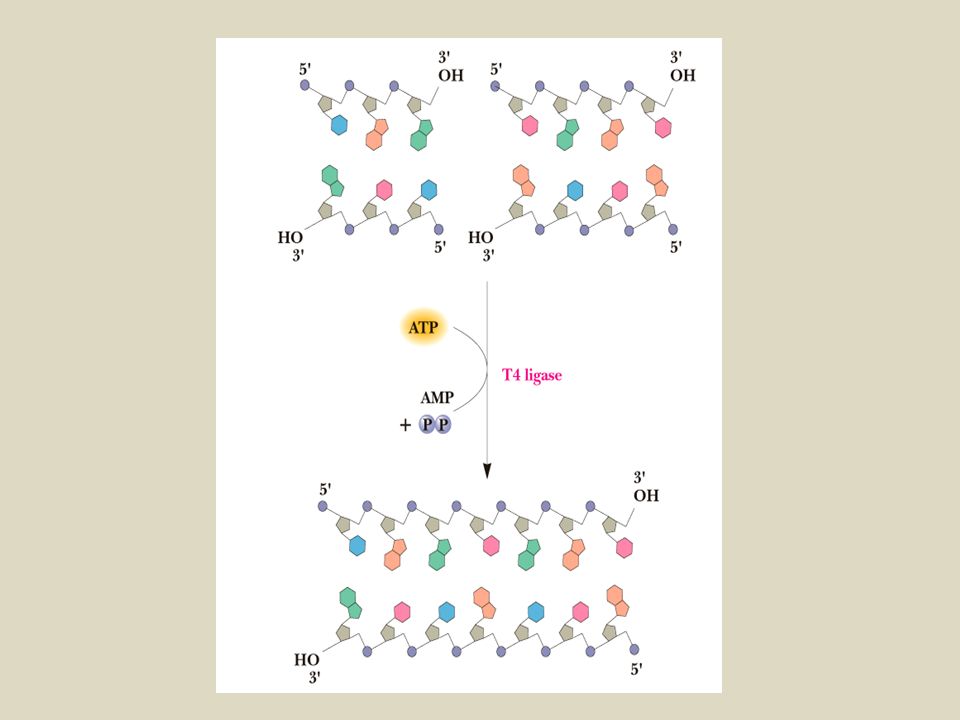
Le DNA ligasi sono enzimi capaci di legare covalentemente due molecole di DNA adiacenti con estremità libere

32
DNA RICOMBINANTE: due tratti di DNA che in natura non sono
adiacenti, vengono uniti in provetta. Le proprietà degli enzimi di restrizione sono state essenziali per la tecnica del DNA ricombinante. I due tratti da unire vengono tagliati con uno stesso enzima di restrizione. EcoRI GAATTC CTTAAG G AATTC CTTAA G

33
Le estremità coesive prodotte da uno stesso enzima possono appaiarsi.
G CTTAA AATTC GAATTC CTTAAG

34
Infine la DNA ligasi viene utilizzata per legare covalentemente le due molecole di DNA
G AATTC CTTAA G 5’ 3’ OH P 5’ 3’ G CTTAA OH P AATTC DNA LIGASI ATP GAATTC CTTAAG 5’ 3’

35
Solo estremità compatibili possono essere legate efficientemente

36
INSERIMENTO DI UN TRATTO DI DNA ESOGENO IN UN VETTORE DI CLONAGGIO

37
CLONAGGIO: - LIGASI - TRASFORMAZIONE - SELEZIONE - CRESCITA COLONIE

38
BATTERICA DI INTERESSE
IDENTIFICAZIONE DELLA COLONIA BATTERICA DI INTERESSE

39
Cosa può succedere in una reazione ligasica ?
Il vettore si richiude su se stesso senza legarsi con l’inserto Il vettore si lega con l’inserto

40
L’enzima beta-galattosidasi può essere utilizzato per discriminare le colonie che contengono un plasmide con inserto da quelle che contengono un plasmide senza inserto

41
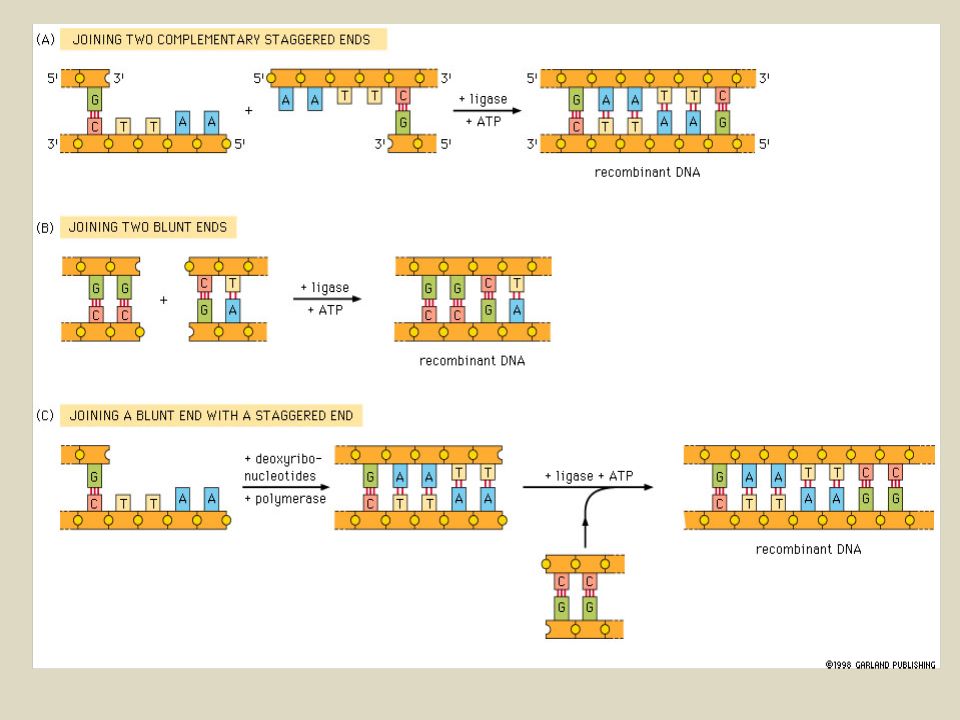
ALCUNI ENZIMI PRODUCONO ESTREMITA’ TRONCHE (BLUNT)
ESTREMITA’ BLUNT POSSONO ESSERE LEGATE A ESTREMITA’ BLUNT PRODOTTE DA QUALSIASI ALTRO ENZIMA, NON CI SONO LE LIMITAZIONI IMPOSTE DALLA COMPLEMENTARIETA’ DELLE BASI DELLE CODINE APPICCICOSE. SVANTAGGIO: LIGASI DI ESTREMITA’ BLUNT E’ MENO EFFICIENTE PECHE’ NON SI HA APPAIAMENTO TRA CODINE A SINGOLO FILAMENTO. CCC/GGG GGG/CCC GAT/ATC CTA/TAG EcoRV SmaI CCC GGG ATC TAG LA SEQUENZA IBRIDA CHE SI FORMA NON E’ RICONOSCIUTA DA NESSUNO DEI DUE ENZIMI.

42
QUANDO E’ NECESSARIO UNIRE DUE ESTREMITA’ INCOMPATIBILI????
E’ POSSIBILE CONVERTIRE LE ESTREMITA’ STICKY IN ESTREMITA’ BLUNT Estremita’ 5’ protruding: 5’ 3’ 5’ 3’ L’estremità 3’ del filamento viene utilizzata come innesco dalla DNA POLIMERASI. Si utilizza il frammento Klenow della DNA polimerasi I di E. coli (DNA POLIMERERASI intera ha azione 5’esonucleasica). Estremità 5’ o 3’ protruding: 5’ 3’ 3’ 5’ Si utilizza la nucleasi S1: nucleasi che funziona su singolo filamento.
. Estremità 5’ o 3’ protruding: 5’ 3’ 3’ 5’ Si utilizza la nucleasi S1: nucleasi che funziona su singolo filamento.")
43
Procedura di selezione
Selezione dei cloni Bisogna individuare il clone batterico che contiene il gene di interesse Occorre inserire dei marcatori genici nel vettore in cui è presente il gene di interesse Procedura di selezione Screening bianco-blu

44
Rappresentazione schematica del clonaggio (I)
DNA SORGENTE VETTORE DI CLONAGGIO 1 DNA BERSAGLIO frammentazione enzimatica linearizzazione enzimatica 2 3 congiungimento DNA bersaglio- vettore di clonazione

45
introduzione del DNA nella
cellula ospite (trasformazione) e isolamento delle cellule con il gene clonato cellula ospite proteina codificata dal gene clonato Produzione della proteina tutto ciò è possibile… grazie alla scoperta delle endonucleasi di restrizione
 e isolamento delle cellule con il gene clonato. cellula ospite. proteina codificata. dal gene clonato. Produzione della proteina. tutto ciò è possibile… grazie alla scoperta delle endonucleasi di restrizione.")
46
Le Endonucleasi di Restrizione (I)
Enzimi batterici: endonucleasi di TIPO II: Riconoscono e tagliano corte sequenze di DNA (4-8bp); le sequenze riconosciute sono palindromiche, cioè risultano ugluali se lette su ogni filamento nella stessa direzione il taglio produce estremità piatte o coesive Esempio di una delle prime endonucleasi di tipo II meglio caratterizzate: EcoRI genere: Escherichia Ordine di caratterizzazione di diverse endonucleasi appartenente allo stesso organismo (I) specie: Coli ceppo: R 5’- GAATTC - 3’ 3’- CTTAAG – 5’ SITO DI RICONOSCIMENTO EcoRI Il taglio genera 4 basi sporgenti in singolo filamento 5’- G AATTC - 3’ 3’- CTTAA G – 5’ OH P PRODOTTI FORMAZIONE DI ESTREMITÀ COESIVE
; le sequenze riconosciute sono palindromiche, cioè risultano ugluali se lette su ogni filamento nella stessa direzione. il taglio produce estremità piatte o coesive. Esempio di una delle prime endonucleasi di tipo II meglio caratterizzate: EcoRI. genere: Escherichia. Ordine di caratterizzazione di diverse endonucleasi appartenente allo stesso organismo (I) specie: Coli. ceppo: R. 5’- GAATTC - 3’ 3’- CTTAAG – 5’ SITO DI. RICONOSCIMENTO. EcoRI. Il taglio genera 4 basi sporgenti in singolo filamento. 5’- G AATTC - 3’ 3’- CTTAA G – 5’ OH. P. PRODOTTI. FORMAZIONE. DI ESTREMITÀ. COESIVE.")
47
Le Endonucleasi di Restrizione (II)
Enzima Sito di riconoscimento EcoR I G A A T T C C T T A A G BamH I G G A T C C C C T A G G Hind III A A G C T T T T C G A A KPN I G G T A C C C C A T G G NOT I G C G G C C G C C G C C G G C G EcoR V G A T A T C C T A T A G Xho I C T C G A G G A G C T C Isoschizomeri Sebbene gli enzimi di restrizione isolati siano oltre 3500, le sequenze bersaglio che possono essere tagliate sono molte di meno (appena più di duecento). E’ evidente che molti enzimi isolati da batteri diversi hanno la stessa specificità di sequenza,sono cioè isoschizomeri.
. E’ evidente che molti enzimi isolati da batteri diversi hanno la stessa specificità di sequenza,sono cioè isoschizomeri.")
48
Esempio: HaeIII (da Haemophilus parainfluenzae)
Estremità piatte Esempio: HaeIII (da Haemophilus parainfluenzae) La sequenza di DNA in doppia elica: 5’-NNNNGGCCNNNN-3’ 3’-NNNNCCGGNNNN-5’ viene tagliata nel seguente modo: 5’-NNNNGG pCCNNNN-3’ 3’-NNNNCCp GGNNNN-5’ Estremità coesive Esempio: EcoRI La sequenza di DNA in doppia elica: 5’-NNNNGAATTCNNNN-3’ 3’-NNNNCTTAAGNNNN-5’ viene tagliata nel seguente modo: 5’-NNNNG pAATTCNNNN-3’ 3’-NNNNCTTAAp GNNNN-5’ In questo caso il taglio è perfettamente simmetrico. Le estremità piatte sono saldate in modo inefficiente dalla DNA ligasi, perché non restano associate a lungo. Tutte le estremità generate dagli enzimi che operano un taglio sfalsato, siano esse sporgenti in 5’ o in 3’, sono “appiccicose”, cioè possono formare ponti idrogeno tra le due code a filamento singolo complementari.
 La sequenza di DNA in doppia elica: 5’-NNNNGGCCNNNN-3’ 3’-NNNNCCGGNNNN-5’ viene tagliata nel seguente modo: 5’-NNNNGG pCCNNNN-3’ 3’-NNNNCCp GGNNNN-5’ Estremità coesive. Esempio: EcoRI. La sequenza di DNA in doppia elica: 5’-NNNNGAATTCNNNN-3’ 3’-NNNNCTTAAGNNNN-5’ viene tagliata nel seguente modo: 5’-NNNNG pAATTCNNNN-3’ 3’-NNNNCTTAAp GNNNN-5’ In questo caso il taglio è perfettamente simmetrico. Le estremità piatte sono saldate in modo inefficiente dalla DNA ligasi, perché non restano associate a lungo. Tutte le estremità generate dagli enzimi che operano un taglio sfalsato, siano esse sporgenti in 5’ o in 3’, sono appiccicose , cioè possono formare ponti idrogeno tra le due code a filamento singolo complementari.")
49
Rappresentazione schematica del clonaggio (II) - estremità coesive
annealing reannealing Tutti questi appaiamenti (annealing-reannealing) sono transitori a causa della debolezza dei legami a idrogeno tra il numero ridotto di basi a livello delle estremità coesive. Questi legami vengono stabilizzati grazie all’uso dell’enzima DNA LIGASI Enzima T4 DNA ligasi (isolato dal batteriofago T4) forma legami covalenti tra il gruppo fosfato 5’ di una estremità e il gruppo 3’ OH della catena adiacente. L’enzima lavora a 10°C e richiede ATP.
 sono transitori a causa della debolezza dei legami a idrogeno tra il numero ridotto di basi a livello delle estremità coesive. Questi legami vengono stabilizzati grazie all’uso dell’enzima DNA LIGASI. Enzima T4 DNA ligasi (isolato dal batteriofago T4) forma legami covalenti tra il gruppo fosfato 5’ di una estremità e il gruppo 3’ OH della catena adiacente. L’enzima lavora a 10°C e richiede ATP.")
50
Rappresentazione schematica del clonaggio (III) - estremità coesive
 - estremità coesive")
51
Trattamento del DNA con estremità piatte
La ligazione di frammenti con estremità piatte non è efficiente quanto quella di frammenti con estremità coesive, perciò prima di unire il DNA con il vettore di clonaggio è necessario eseguire alcune procedure aggiuntive: AGGIUNTA DEL LINKER plasmide DNA a estremità piatte 5’ pGGGATCCC 3 CCCTAGGG linker Unione del linker al DNA con estramità piatte mediante DNA ligasi 5’ pGGGATCCC 3’ CCCTAGGG GGGATCCC CCCTAGGG Taglio con BamHI CTAGp 5’pGATC 3’ IN ALTRI CASI L POTO DEL LINKER SI PUò AGGIUNGERE UN ADATTAORE CHE HA GIA ESTREMITà COESIVE COSI DA NON PASSARE PER LA DIGESTIONE ENZIMATICA 5’ pGATCCC 3’ GG GG CCCTAGp Taglio con BamHI C T A G G G G G G A T C G A T C C C C C C T A G Unione delle estremità coesive mediante DNA ligasi

52
Vettori per il clonaggio (I)
Per clonare una qualsiasi molecola di DNA è necessario inserirla in un vettore per il clonaggio. Questi elementi di DNA possono essere mantenuti e propagati in un organismo ospite in cui il vettore possa replicarsi. Un tipico ospite è Escherichia Coli che cresce e si divide rapidamente. Qualsiasi vettore con un origine di replicazione adatta si replica con successo in E.Coli (assieme al DNA inserito). Quindi, qualsiasi DNA inserito in un vettore può essere amplificato (se ne possono produrre quantità illimitate) e usato per successive analisi Libreria genica: ogni vettore contiene un frammento diverso di DNA estraneo Isolamento di un clone mediante screening della libreria Dire che I vettori che si utilizzano sono diversi e differiscon per complessità, selezione e lunghezza di DNA che si può inserire. In genere I vettori sono stati sviluppati da molecole presenti in natura Amplificazione del singolo clone
. Quindi, qualsiasi DNA inserito in un vettore può essere amplificato (se ne possono produrre quantità illimitate) e usato per successive analisi. Libreria genica: ogni vettore contiene un frammento diverso di DNA estraneo. Isolamento di un clone mediante screening della libreria. Dire che I vettori che si utilizzano sono diversi e differiscon per complessità, selezione e lunghezza di DNA che si può inserire. In genere I vettori sono stati sviluppati da molecole presenti in natura. Amplificazione. del singolo clone.")
53
Vettori per il clonaggio (II)
- SONO STATI SVILUPPATI DA MOLECOLE PRESENTI IN NATURA (PLASMIDI BATTERICI, BATTERIOFAGI) O COMBINAZIONE DEGLI ELEMENTI CHE LI COMPONGONO (COSMIDI). Plasmidi (i) DNA extracromosomico circolare di piccole dimensioni presente in molti batteri che conferisce resistenza a antibiotici, capacità di metabolizzare substrati e capacità di coniugazione. anni ’70: alcuni di questi vettori sono stati modificati in laboratorio (digestioni e successive ligazioni) per costruire vettori di clonaggio (es: pBR322 da Bolivar e Rodriguez); un plasmide artificiale è molto più piccolo di quello naturale e viene più facilmente inserito nei batteri nel processo noto come trasformazione; un origine di replicazione del DNA di tipo batterico cosicchè il DNA può essere replicato dalla cellula ospite plasmidi come pBR322 hanno un origine di replicazione rilassata, che non segue cioè la divisione cellulare, in modo che la replicazione del plasmide è molto più frequente di quella cromosomica. In questo modo ogni cellula produce molte molecole di plasmide. Sono stati introdotti due geni che codificano per la resistenza agli antibiotici in vari punti del palsmide ci sono siti singoli di riconoscimento per una serie di enzimi di restrizione. Alcuni di questi siti possono essere inseriti in un gene che codifica per la resistenza ad un antibiotico, cosicchè la presenza di un particolare inserto può essere rivelata dalla perdita di resistenza a quello stesso antibiotico (inattivazione inserzionale).
 O COMBINAZIONE DEGLI ELEMENTI CHE LI COMPONGONO (COSMIDI). Plasmidi (i) DNA extracromosomico circolare di piccole dimensioni presente in molti batteri che conferisce resistenza a antibiotici, capacità di metabolizzare substrati e capacità di coniugazione. anni ’70: alcuni di questi vettori sono stati modificati in laboratorio (digestioni e successive ligazioni) per costruire vettori di clonaggio (es: pBR322 da Bolivar e Rodriguez); un plasmide artificiale è molto più piccolo di quello naturale e viene più facilmente inserito nei batteri nel processo noto come trasformazione; un origine di replicazione del DNA di tipo batterico cosicchè il DNA può essere replicato dalla cellula ospite. plasmidi come pBR322 hanno un origine di replicazione rilassata, che non segue cioè la divisione cellulare, in modo che la replicazione del plasmide è molto più frequente di quella cromosomica. In questo modo ogni cellula produce molte molecole di plasmide. Sono stati introdotti due geni che codificano per la resistenza agli antibiotici. in vari punti del palsmide ci sono siti singoli di riconoscimento per una serie di enzimi di restrizione. Alcuni di questi siti possono essere inseriti in un gene che codifica per la resistenza ad un antibiotico, cosicchè la presenza di un particolare inserto può essere rivelata dalla perdita di resistenza a quello stesso antibiotico (inattivazione inserzionale).")
54
Vettori per il clonaggio (III)
pBR322 Esempio di mutagenesi inserzionale (singolo taglio con BamHI)
")
55
Plasmidi (ii) : pUC gene per la resistenza alla tetraciclina;
origine di replicazione per E.Coli; Multiple Cloning Site (MCS) o polylinker: siti di restrizione unici più comuni; MCS è parte del gene lacZ che codifica per la b-galattosidasi;
 o polylinker: siti di restrizione unici più comuni; MCS è parte del gene lacZ che codifica per la b-galattosidasi;")
56
Plasmidi (iii) : pUC MCS Vettori non ricombinanti (senza inserto)
Quando si trasformano cellule di E.Coli con il plasmide pUC, il gene può essere attivato aggiungnedo nel terreno l’induttore isopropil-b-D-tiogalattopiranoside (IPTG), la cui presenza induce l’espressione della b-galattosidasi. L’ezima funzionale idrolizza una sostanza incolore detta 5-bromo-4-cloro-3-indolil-b-galattopiranoside (X-Gal) producendo un composto blu insolubile. Vettori non ricombinanti (senza inserto) MCS GENE PER LA b-GALATTOSIDASI X-Gal idrolizzato: placca blu INDUZIONE CON IPTG 2. Vettori ricombinanti (senza inserto) INSERIMENTO DEL DNA NEL MCS CON INTERRUZIONE DEL GENE b-GAL INDUZIONE CON IPTG GENE INSERITO X-Gal NON idrolizzato: placca bianca
, la cui presenza induce l’espressione della b-galattosidasi. L’ezima funzionale idrolizza una sostanza incolore detta 5-bromo-4-cloro-3-indolil-b-galattopiranoside (X-Gal) producendo un composto blu insolubile. Vettori non ricombinanti (senza inserto) MCS. GENE PER LA b-GALATTOSIDASI. X-Gal idrolizzato: placca blu. INDUZIONE CON IPTG. 2. Vettori ricombinanti (senza inserto) INSERIMENTO DEL DNA NEL MCS CON INTERRUZIONE DEL GENE b-GAL. INDUZIONE CON IPTG. GENE INSERITO. X-Gal NON idrolizzato: placca bianca.")
57
Procedura di selezione
Screening bianco-blu Il plasmide (pUC19) contiene due marcatori genetici utili per la selezione: gene amp e gene lac Z Il DNA esogeno viene inserito nel plasmide, dove inattiva il gene lac Z Il plasmide ricombinante viene introdotto nel batterio che diventa ampicillina resistente I batteri vengono seminati su piastre contenenti ampicillina e il substrato per la beta-galattossidasi La crescita di colonie bianche indica la presenza del DNA esogeno. Le colonie blu contengono il gene lacZ e quindi il DNA esogeno non si è inserito
 contiene due marcatori genetici utili per la selezione: gene amp e gene lac Z. Il DNA esogeno viene inserito nel plasmide, dove inattiva il gene lac Z. Il plasmide ricombinante viene introdotto nel batterio che diventa ampicillina resistente. I batteri vengono seminati su piastre contenenti ampicillina e il substrato per la beta-galattossidasi. La crescita di colonie bianche indica la presenza del DNA esogeno. Le colonie blu contengono il gene lacZ e quindi il DNA esogeno non si è inserito.")
58
Vettori derivati da virus (I)
Per inserti lunghi (più di 5kb) si usano vettori derivanti dal batteriofago l in quanto offrono una efficienza di clonaggio superiore di circa 16 volte rispetto ai vettori plasmidici; il batteriofago l è un fago con DNA lineare a doppia elica, lungo circa 49 kb in grado di infettare E.Coli, inserendogli il proprio DNA attraverso la membrana cellulare il DNA del fago l può replicarsi secondo due modalità: si integra nel cromosoma di E. Coli dove resta quiesciente fino a quando un segnale ne promuove l’escissione (ciclo lisogenico) si replica aapena infetta la cellula, sintetizza le proteine della testa e della coda, si formano nuove particelle fagiche (ciclo litico) PER GLI INSERTI LUNGHI (GIA QUELLI DI 5KB) AUMENATO TANTO LE DIMENSIONI DEI PLASMIDI AL PUNTO CHE L’EFFICIENZA DELLA TRASFORMAZIONE DELLE CELLULE BATTERIUCHE DIMINUISCE MOLTO
 si usano vettori derivanti dal batteriofago l in quanto offrono una efficienza di clonaggio superiore di circa 16 volte rispetto ai vettori plasmidici; il batteriofago l è un fago con DNA lineare a doppia elica, lungo circa 49 kb in grado di infettare E.Coli, inserendogli il proprio DNA attraverso la membrana cellulare. il DNA del fago l può replicarsi secondo due modalità: si integra nel cromosoma di E. Coli dove resta quiesciente fino a quando un segnale ne promuove l’escissione (ciclo lisogenico) si replica aapena infetta la cellula, sintetizza le proteine della testa e della coda, si formano nuove particelle fagiche (ciclo litico) PER GLI INSERTI LUNGHI (GIA QUELLI DI 5KB) AUMENATO TANTO LE DIMENSIONI DEI PLASMIDI AL PUNTO CHE L’EFFICIENZA DELLA TRASFORMAZIONE DELLE CELLULE BATTERIUCHE DIMINUISCE MOLTO.")
59
Vettori derivati da virus (II)
coda DNA testa I fagi sono stati utilizzati per la costruzione di librerie geniche perchè hanno permesso di introdurre frammenti più grandi di DNA rispetto ai plasmidi. Il DNA del batteriofago l è lungo circa 50 kb, approssimativamente 20 dei quali risultano indispensabili agli eventi di integrazione-escissione (I/E). Per formare le genoteche si è ragionato sulla possibilità di sostituire queste 20 kb con altrettante kb di DNA clonato. Una simile molecola di DNA si potrebbe perpetuare sotto forma di batteriofago l “ricombinante” tramite cicli litici forzosi.
. Per formare le genoteche si è ragionato sulla possibilità di sostituire queste 20 kb con altrettante kb di DNA clonato. Una simile molecola di DNA si potrebbe perpetuare sotto forma di batteriofago l ricombinante tramite cicli litici forzosi.")
60
Cosmidi Vettori di clonaggio Vector system Host cell
Possono trasportare 40 kb di DNA clonato e possono essere mantenuti come plasmidi in E. Coli. Vettori di clonaggio Vector system Host cell Insert capacity (kb) Plasmid E. coli 0.1-10 Bacteriophage l 10-20 Cosmid 35-45 Bacteriophage P1 80-100 BAC (bacterial artificial chromosome) 50-300 P1 bacteriophage-derived AC YAC Yeast 100-2,000
 Plasmid. E. coli Bacteriophage l Cosmid Bacteriophage P BAC (bacterial artificial chromosome) P1 bacteriophage-derived AC YAC. Yeast ,000.")
61
La trasformazione (I) introduzione del DNA nella
cellula ospite (trasformazione) e isolamento delle cellule con il gene clonato Produzione della proteina dal gene clonato proteina codificata dal gene clonato
 e isolamento delle cellule con il gene clonato. Produzione della proteina. dal gene clonato. proteina codificata. dal gene clonato.")
62
La trasformazione (II)
TRASFORMAZIONE: assunzione di DNA da parte di un organismo 1970: Mandel and Higa osservarono che batteri, trattati con soluzioni di CaCl2 e riscaldati velocemente, potevano essere trasformati con DNA del batteriofago λ quindi questo trattamento conferiva ai batteri un transiente stato di “competenza” COMPETENZA: capacità di assumere DNA = numero di colonie positive/microgrammo di DNA trasformato Nel corso degli anni sono stati fatti diversi tentativi per migliorare l’efficienza di trasformazione. Le due tecniche maggiormente utilizzate oggi sono: Trasformazione per CaCl2 Elettroporazione

63
Trasformazione Per Elettroporazione
Le cellule batteriche unite al DNA vengono sottoposte ad una veloce scarica elettrica che permette la formazione di “elettropori” che favoriscono l’entrata di DNA. Competenza: cellule trasformate /μg di DNA

64
Trasformazione in CaCl2
Crescita di un ceppo E.Coli in terreno ricco di nutrienti. Le cellule si raccolgono quando sono in fase logaritmica

65
I cationi divalenti schermano le cariche negative sul DNA (dei gruppi fosfato)
La combinazione di bassa temperatura e di cationi divalenti può aiutare a cristallizzare regioni della membrana rendendo più accessibili i canali di assunzione del DNA

66
Sommario delle fasi importanti nell clonaggio del DNA
Vettore plasmidico e DNA esogeno da inserire devono avere estremità compatibili. I due “pezzi” di DNA sono fusi creando molecole di DNA ricombinante; Introduzione del DNA in cellule batteriche ospiti idonee (trasformazione) Selezione delle cellule resistenti all’antibiotico e selezione dei cloni che hanno assunto il DNA ricombinante
 Selezione delle cellule resistenti all’antibiotico e selezione dei cloni che hanno assunto il DNA ricombinante.")
67
esempio di clonaggio gene di interesse BamHI EcoRI VETTORE I

68
esempio di clonaggio …Come si procede ???... gene di interesse Not1
BglII VETTORE I …Come si procede ???... se decido di clonare il gene di interesse BamHI-EcoRI devo essere sicura che gli stessi enzimi non tagliano all’interno della mia sequenza genica; Devo disegnare due oligo, uno frw con la sequenza che possiede il sito di riconoscimento per l’enzima BamHI e uno reverse che contiene il sito di riconoscimento per l’enzima EcoRI Amplificazione per PCR della sequenza genica di interesse con gli oligo disegnati; Purificazione della PCR preparativa Digestione con gli enzimi BamHI e EcoRI per la formazione delle estremità coesive Digestione del vettore pOTB7 con BamH1 e EcoRI Ligazione con l’ezima T4 DNA ligasi

69
1 atggagatgg aaaaggagtt cgagcagatc gacaagtccg ggagctgggc ggccatttac
se decido di clonare il gene di interesse BamHI-EcoRI devo essere sicura che gli stessi enzimi non tagliano all’interno della mia sequenza genica; 1 atggagatgg aaaaggagtt cgagcagatc gacaagtccg ggagctgggc ggccatttac 61 caggatatcc gacatgaagc cagtgacttc ccatgtagag tggccaagct tcctaagaac 121 aaaaaccgaa ataggtacag agacgtcagt ccctttgacc atagtcggat taaactacat 181 caagaagata atgactatat caacgctagt ttgataaaaa tggaagaagc ccaaaggagt 241 tacattctta cccagggccc tttgcctaac acatgcggtc acttttggga gatggtgtgg 301 gagcagaaaa gcaggggtgt cgtcatgctc aacagagtga tggagaaagg ttcgttaaaa 361 tgcgcacaat actggccaca aaaagaagaa aaagagatga tctttgaaga cacaaatttg 421 aaattaacat tgatctctga agatatcaag tcatattata cagtgcgaca gctagaattg 481 gaaaacctta caacccaaga aactcgagag atcttacatt tccactatac cacatggcct 541 gactttggag tccctgaatc accagcctca ttcttgaact ttcttttcaa agtccgagag 601 tcagggtcac tcagcccgga gcacgggccc gttgtggtgc actgcagtgc aggcatcggc 661 aggtctggaa ccttctgtct ggctgatacc tgcctcttgc tgatggacaa gaggaaagac 721 ccttcttccg ttgatatcaa gaaagtgctg ttagaaatga ggaagtttcg gatggggctg 781 atccagacag ccgaccagct gcgcttctcc tacctggctg tgatcgaagg tgccaaattc 841 atcatggggg actcttccgt gcaggatcag tggaaggagc tttcccacga ggacctggag 901 cccccacccg agcatatccc cccacctccc cggccaccca aacgaatcct ggagccacac 961 aatgggaaat gcagggagtt cttcccaaat caccagtggg tgaaggaaga gacccaggag 1021 gataaagact gccccatcaa ggaagaaaaa ggaagcccct taaatgccgc accctacggc 1081 atcgaaagca tgagtcaaga cactgaagtt agaagtcggg tcgtgggggg aagtcttcga 1141 ggtgcccagg ctgcctcccc agccaaaggg gagccgtcac tgcccgagaa ggacgaggac 1201 catgcactga gttactggaa gcccttcctg gtcaacatgt gcgtggctac ggtcctcacg 1261 gccggcgctt acctctgcta caggttcctg ttcaacagca acacatag Programma che permette di fare un’analisi di restrizione della sequenza

71
CTAGCGGATCCGGCCCGTCATGGAGATGGAA
2. Devo disegnare due oligo, uno frw con la sequenza che possiede il sito di riconoscimento per l’enzima BamHI e uno reverse che contiene il sito di riconoscimento per l’enzima EcoRI ggcccgtcatggagatggaaaaggagttcgagcagatcgacaagtccgggagctgggcggccatttac gccggcgcttacctctgctacaggttcctgttcaacagcaacacatag Costruzione dell’oligo frw Codina di 5-6 nt - sequenza riconosciuta dall’enzima di restrizione (BamHI) – sequenza compresa di atg (codone di inizio). L’oligo deve essere lungo circa nt CTAGCGGATCCGGCCCGTCATGGAGATGGAA Costruzione dell’oligo reverse Sequenza fino al codone di stop (stop incluso)- sequenza riconosciuta dall’enzima di restrizione- e codina di 5-6 nt. Fare il reverse complement TTCCTGTTCAACAGCAACACATAGGAATTCCTAGC GCTAGGAATTCCTATGTGTTGCTGTTGAACAGGAA
 – sequenza compresa di atg (codone di inizio). L’oligo deve essere lungo circa nt. CTAGCGGATCCGGCCCGTCATGGAGATGGAA. Costruzione dell’oligo reverse. Sequenza fino al codone di stop (stop incluso)- sequenza riconosciuta dall’enzima di restrizione- e codina di 5-6 nt. Fare il reverse complement. TTCCTGTTCAACAGCAACACATAGGAATTCCTAGC. GCTAGGAATTCCTATGTGTTGCTGTTGAACAGGAA.")
72
Purificazione della PCR preparativa
CTAGCGGATCCGGCCCGTCATGGAGATGGAA CCGGGCAGTACCTCTACCTTTTCCTC GGCCCGTCATGGAGATGGAAAAGGAGTTCGAGCAGATCGACAAGTCCGGGAGCTGGGCGGCCATTTAC CGGCCGCGAATGGAGACGATGTCCAAGGACAAGTTGTCGTTGTGTATC GCCGGCGCTTACCTCTGCTACAGGTTCCTGTTCAACAGCAACACATAG AAGGACAAGTTTGTCGTTGTGTATCCTTAAGGATCG Amplificazione per PCR della sequenza genica di interesse con gli oligo disegnati; Purificazione della PCR preparativa Digestione con gli enzimi BamHI e EcoRI per la formazione delle estremità coesive Digestione del vettore pOTB7 con BamH1 e EcoRI Ligazione con l’ezima T4 DNA ligasi

73
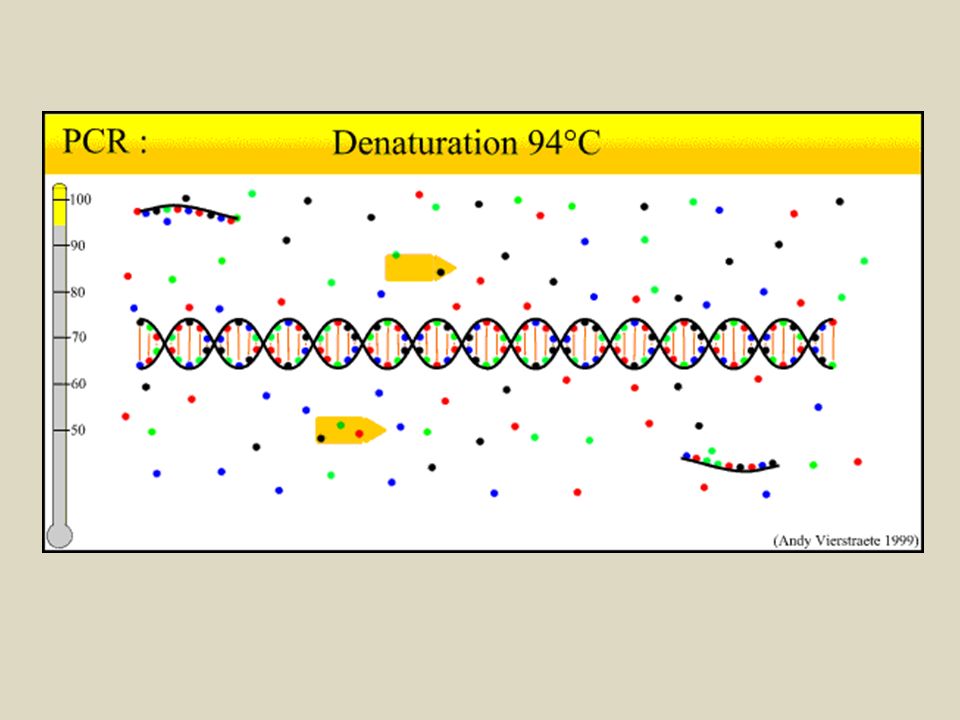
Reazione a catena della polimerasi, PCR
E’ una tecnica che consente di amplificare piccole quantità di DNA DNA polimerasi di Thermus aquaticus

74
PCR Trenta cicli incrementano la quantità di DNA di oltre un miliardo di volte!!!

Presentazioni simili