Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
ENZIMI - (quasi) Tutti gli enzimi sono proteine
Sono catalizzatori biologici non vengono alterati dalla reazione che catalizzano ACCELERANO reazioni chimiche fino a 1020 volte!!! (efficienza catalitica) Nota:accelerazione da catalizzatori non enzimatici: Rendono più moderate le condizioni di reazione altrimenti drastiche per pH, temperatura, pressione…
 Nota:accelerazione da catalizzatori non enzimatici: Rendono più moderate le condizioni di reazione altrimenti drastiche per pH, temperatura, pressione…")
2
Gli enzimi sono catalizzatori biologici
specificità di substrato substrati = reagenti su cui gli enzimi agiscono specificità di reazione (assenza di prodotti secondari) Possono essere regolati in funzione delle specifiche esigenze
 Possono essere regolati in funzione delle specifiche esigenze.")
3
Che cosa gli enzimi NON fanno:
NON cambiano l’equilibrio di una reazione - NON rendono possibile una reazione termodinamicamente impossibile

4
Gli enzimi possono avere dimensioni variabili
Solo una piccola parte della molecola è coinvolta nella catalisi (sito catalitico o sito attivo), ma…tutta la molecola è importante per la funzione Alterazioni a siti distanti dal sito catalitico (anche in un solo aa!!!) possono ridurre o annullare l’attività dell’enzima!!! Per svolgere la sua funzione l’enzima DEVE legare il substrato Formazione di un complesso enzima-substrato (ES)
, ma…tutta la molecola è importante per la funzione. Alterazioni a siti distanti dal sito catalitico (anche in un solo aa!!!) possono ridurre o annullare l’attività dell’enzima!!! Per svolgere la sua funzione l’enzima DEVE legare il substrato. Formazione di un complesso enzima-substrato (ES)")
5
COMPLEMENTARIETA’ GEOMETRICA COMPLEMENTARIETA’ ELETTRONICA
Il sito attivo forma una superficie tridimensionale complementare al substrato COMPLEMENTARIETA’ GEOMETRICA COMPLEMENTARIETA’ ELETTRONICA STEREOSPECIFICITA’ L’enzima lega il substrato mediante forze di van der Waals, ponti idrogeno, interazioni idrofobiche…

6
Chimotripsina: proteasi
Specificità: idrolisi di legami peptidici adiacenti a residui aromatici o con lunghe catene laterali idrofobiche Complesso ES

7
Interazione enzima-substrato
Chiave-serratura Emil Fisher, 1890 Non tiene conto della flessibilità conformazionale delle proteine Adattamento indotto Daniel Koshland, 1958 Il sito attivo assume una forma complementare a quella del substrato solo DOPO che il substrato si è legato

8
Esochinasi Glucosio + ATP glucosio 6-fosfato + ADP

9
Nomenclatura degli enzimi
Nome del substrato su cui agiscono (o del tipo di reazione che catalizzano) + suffisso –asi Es: - ureasi - Alcool deidrogenasi - glucosio 6-fosfato isomerasi - DNA ligasi - esochinasi
 + suffisso –asi. Es: - ureasi. - Alcool deidrogenasi. - glucosio 6-fosfato isomerasi. - DNA ligasi. - esochinasi.")
10
Regole per la classificazione degli enzimi
International Union of Biochemistry and Molecular Biology (IUBMB) Gli enzimi vengono classificati in base al tipo di reazione chimica che catalizzano
 Gli enzimi vengono classificati in base al tipo di reazione chimica che catalizzano.")
11
Classificazione Tipo di reazione catalizzata
1. Ossidoreduttasi Reazioni di ossidoriduzione (deidrogenasi, ossidasi, perossidasi, reduttasi) 2. Trasferasi trasferimento di gruppi funzionali 3. Idrolasi idrolisi Liasi lisi del substrato con formazione di doppi legami (reazioni di eliminazione) (nella reazione inversa -reazione di addizione - agiscono da sintasi) 5. Isomerasi isomerizzazione 6. Ligasi formazione di legami accoppiata all’idrolisi di ATP (anche denominate “sintetasi”)
 2. Trasferasi trasferimento di gruppi funzionali. 3. Idrolasi idrolisi. Liasi lisi del substrato con formazione di doppi. legami (reazioni di eliminazione) (nella reazione inversa -reazione di. addizione - agiscono da sintasi) 5. Isomerasi isomerizzazione. 6. Ligasi formazione di legami accoppiata all’idrolisi. di ATP. (anche denominate sintetasi )")
12
A ciascun enzima vengono assegnati due nomi e un numero
di classificazione: Nome corrente (es. esochinasi) Nome sistematico (es: ATP glucosio fosfotransferasi) Numero di classificazione (es: EC ) Enzyme Commission numero di serie classe sottoclasse sotto-sottoclasse
 Nome sistematico (es: ATP glucosio fosfotransferasi) Numero di classificazione (es: EC ) Enzyme Commission. numero di serie. classe. sottoclasse. sotto-sottoclasse.")
13
Di che cosa necessitano gli enzimi per la loro funzione di catalizzatori biologici?
Solo delle catene laterali di specifici amminoacidi nel sito attivo Composti chimici addizionali, i cofattori ioni inorganici, es. Fe2+, Cu2+, K+ , Mg2+, Mn2+, Ni2+, Zn2+ …. complesse molecole organiche e metallorganiche, i coenzimi

14
Il coenzima legato saldamente all’enzima viene detto gruppo prostetico
cofattore Oloenzima (attivo) Apoenzima (inattivo) Il coenzima legato saldamente all’enzima viene detto gruppo prostetico
 Apoenzima. (inattivo) Il coenzima legato saldamente all’enzima viene detto gruppo prostetico.")
15
I coenzimi operano come trasportatori di gruppi funzionali o elettroni
molti coenzimi derivano da vitamine NAD: Trasferimento di elettroni (ioni idruro H-) (niacina ) FAD: Trasferimento di elettroni (riboflavina: vitamina B2) Piridossal fosfato: Trasferimento gruppi amminici (piridossina: vitamina B6) Tiamina pirofosfato: Trasferimento gruppi aldeidici (tiamina: vitamina B1) Biotina: trasferimento CO2 (vitamina B8) Coenzima A e Lipoato: trasferimento di gruppi acilici (pantotenato: vitamina B5)
 (niacina ) FAD: Trasferimento di elettroni. (riboflavina: vitamina B2) Piridossal fosfato: Trasferimento gruppi amminici. (piridossina: vitamina B6) Tiamina pirofosfato: Trasferimento gruppi aldeidici. (tiamina: vitamina B1) Biotina: trasferimento CO2. (vitamina B8) Coenzima A e Lipoato: trasferimento di gruppi acilici. (pantotenato: vitamina B5)")
16
Il meccanismo d’azione degli enzimi può essere trattato da due punti di vista:
analizzando i cambiamenti energetici che si verificano nel corso della reazione esaminando il modo in cui il sito attivo favorisce la catalisi dal punto di vista chimico Energetica Chimica

17
La reazione richiede energia Reazione termodinamicamente
Cambiamenti energetici che si verificano durante una reazione ∆G > 0 La reazione richiede energia DG < 0 Reazione termodinamicamente favorita ∆G = 0 La reazione è all’equilibrio DGo’ = variazione di energia libera in condizioni standard Concentrazione: 1 M di reagenti e prodotti Pressione: 1 Atm Temperatura : 298 °K (25 °C) pH 7
 pH 7.")
18
Cambiamenti energetici che si verificano durante una reazione
Stato di transizione = stato ad alta energia che deve essere raggiunto perché S possa essere convertito in P Energia libera di attivazione: barriera energetica che separa i reagenti dai prodotti

19
…in una reazione non catalizzata solo un piccolo numero di molecole
La velocità della reazione dipende dal superamento della barriera energetica per il raggiungimento dello stato di transizione …in una reazione non catalizzata solo un piccolo numero di molecole possiede energia sufficiente per raggiungere lo stato di transizione

20
I catalizzatori abbassano l’energia di attivazione senza alterare l’equilibrio della reazione
Gli enzimi sono catalizzatori biologici - non vengono consumati durante la reazione - non alterano l’equilibrio della reazione - aumentano la velocità della reazione

21
come fanno gli enzimi ad abbassare l’energia di attivazione?..
…meccanismo???... EFFETTI DI LEGAME energia di legame prossimità e tensione stabilizzazione dello stato di transizione EFFETTI CHIMICI catalisi acido-base catalisi covalente

22
Energia di legame = energia che si libera dall’interazione
Stato di transizione formazione del prodotto rilascio del prodotto Energia di legame = energia che si libera dall’interazione enzima-substrato

23
Il numero di interazioni deboli è massimo allo stato di transizione
EFFETTI DI LEGAME Il numero di interazioni deboli è massimo allo stato di transizione - Massima energia liberata- Le interazioni deboli sono ottimali allo stato di transizione Abbassamento dell’energia di attivazione Stabilizzazione dello stato di transizione

24
cambiamenti conformazionali conseguenti all’interazione
Effetti di tensione cambiamenti conformazionali conseguenti all’interazione stabilizzazione dello stato di transizione L’enzima è complementare allo stato di transizione

25
gruppi funzionali vicini e nel corretto orientamento

26
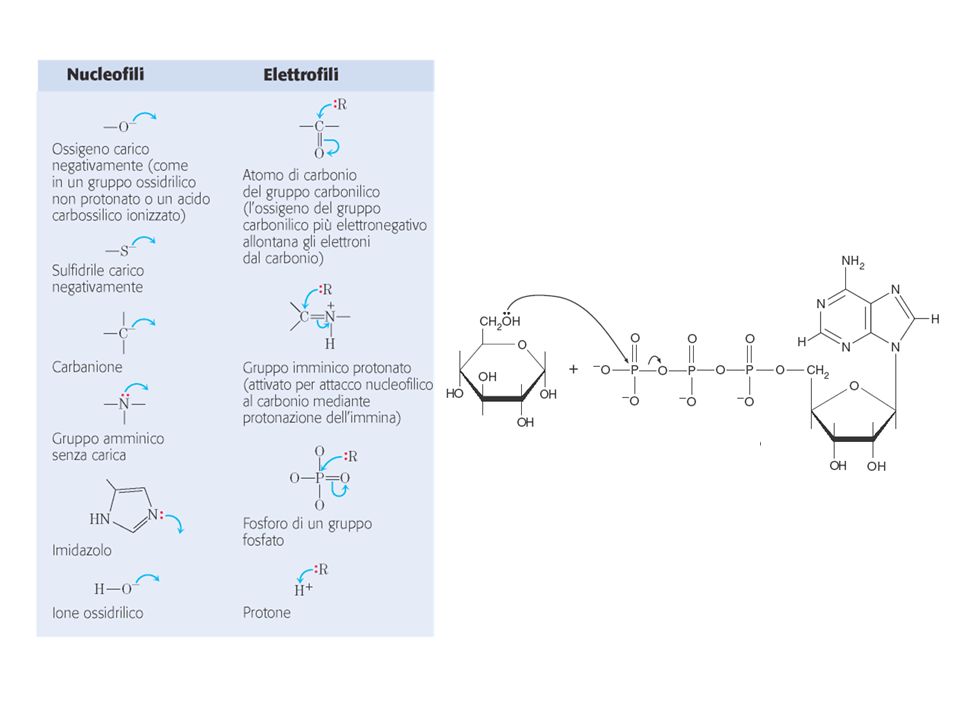
Come leggere un meccanismo di reazione…
Punti = elettroni non condivisi Frecce ricurve = movimento della coppia di elettroni (direzione: nucleofilo elettrofilo) Se la freccia va da un atomo a un altro atomo: si forma un nuovo legame tra i due atomi Se la freccia va da un legame a un atomo: rottura del legame; gli elettroni compaiono come doppietto non condiviso (o carica negativa) sull’atomo indicato dalla freccia Se la freccia va da un legame singolo a un altro legame singolo: si forma un doppio legame .. C C C C C C
 Se la freccia va da un atomo a un altro atomo: si forma un nuovo legame tra i due atomi. Se la freccia va da un legame a un atomo: rottura del legame; gli elettroni compaiono come doppietto non condiviso (o carica negativa) sull’atomo indicato dalla freccia. Se la freccia va da un legame singolo a un altro legame singolo: si forma un doppio legame. .. C C C. C C C.")
28
Catalisi acido-base EFFETTI CHIMICI
Nota: il pKa della catena laterale degli aa può variare in una proteina: effetto dell’ambiente circostante…

29
Catalisi acido-base

30
(Ser, Cys, Glu, Asp, Tyr, His)
EFFETTI CHIMICI Catalisi covalente (Ser, Cys, Glu, Asp, Tyr, His) R1-COOH H2O
 R1-COOH. H2O.")
31
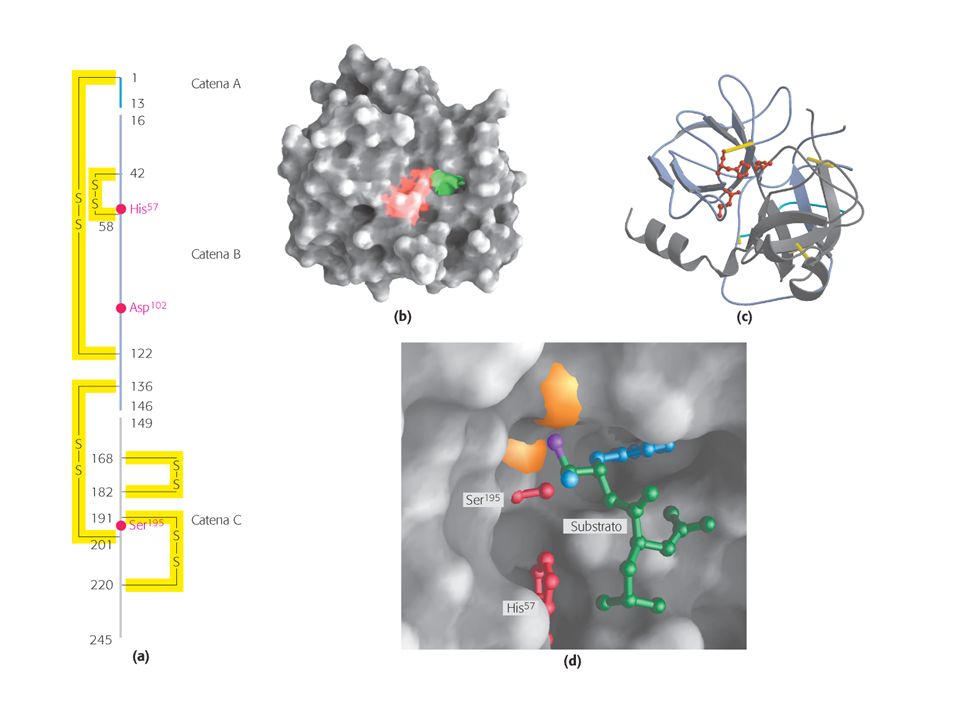
Triade catalitica: Asp – His - Ser
EFFETTI CHIMICI Catalisi covalente Triade catalitica: Asp – His - Ser His57: posiziona correttamente il gruppo OH della Ser e ne aumenta la nucleofilicità Asp102: posiziona correttamente His57 e ne aumenta la capacità di accettare protoni

32
Triade catalitica: Asp – His – Ser
presente anche in altre proteasi Es: tripsina (specificità: Lys, Arg) elastasi (specificità: amminacidi con catene laterali piccole (es: Ala, Ser…) chimotripsina (specificità amminoacidi aromatici, idrofobici voluminosi) Che cosa conferisce specificità?
 elastasi (specificità: amminacidi con catene laterali. piccole (es: Ala, Ser…) chimotripsina (specificità amminoacidi aromatici, idrofobici voluminosi) Che cosa conferisce specificità")
33
Catalisi da ioni metallici
Enzimi che richiedono cofattori Catalisi da ioni metallici - orientamento del substrato nel sito attivo - stabilizzazione dello stato di transizione - ossidoriduzione

34
Enzimi che richiedono cofattori

35
Enzimi che richiedono coenzimi

36
Come studiare il meccanismo d’azione degli enzimi?...
Cinetica Velocità della reazione in funzione di parametri diversi (concentrazione dei reagenti, temperatura, pH…)
")
37
v0 = Vmax [S] KM +[S] Equazione di Michaelis-Menten
Reazione catalizzata da enzimi Effetto della [S] k3 k1 k2 Reazione non catalizzata v0 = Vmax [S] KM +[S] Equazione di Michaelis-Menten Descrive la variazione della velocità di una reazione catalizzata da un enzima al variare della concentrazione di substrato
![v0 = Vmax [S] KM +[S] Equazione di Michaelis-Menten](http://slideplayer.it/slide/10564631/33/images/37/v0+%3D+Vmax+%5BS%5D+KM+%2B%5BS%5D+Equazione+di+Michaelis-Menten.jpg "Reazione catalizzata da enzimi. Effetto della [S] k3. k1. k2. Reazione non catalizzata. v0 = Vmax [S] KM +[S] Equazione di Michaelis-Menten. Descrive la variazione della velocità di una reazione catalizzata da un enzima al variare. della concentrazione di substrato.")
38
Derivazione dell’equazione di Michaelis-Menten
Assunzioni: [S] >> [E] La velocità della reazione inversa è trascurabile Stato stazionario: [ES] costante k3 k1 k2 Quando [S] >>[E], la velocità della reazione è proporzionale alla concentrazione di ES La dissociazione di ES in E + P è la tappa limitante v0 = k3 [ES]

39
v0 = k3 [ES] K1 [E] [S] = k2 [ES] + k3 [ES]
Stato stazionario K1 [E] [S] = k2 [ES] + k3 [ES] K1 [E] [S] = (k2 + k3) [ES] [E] [S] = k2 + k3 [ES] k1 KM [E] [S] = KM [ES] [E] = [Et] – [ES] ([Et] –[ES]) [S] = KM [ES]
![v0 = k3 [ES] K1 [E] [S] = k2 [ES] + k3 [ES]](http://slideplayer.it/slide/10564631/33/images/39/v0+%3D+k3+%5BES%5D+K1+%5BE%5D+%5BS%5D+%3D+k2+%5BES%5D+%2B+k3+%5BES%5D.jpg "Stato stazionario. K1 [E] [S] = k2 [ES] + k3 [ES] K1 [E] [S] = (k2 + k3) [ES] [E] [S] = k2 + k3. [ES] k1. KM. [E] [S] = KM. [ES] [E] = [Et] – [ES] ([Et] –[ES]) [S] = KM. [ES]")
40
v0 = Vmax [S] KM +[S] v0 = k3 [ES] ([Et] –[ES]) [S] = KM [ES]
Stato stazionario k3 k1 k2 v0 = k3 [ES] ([Et] –[ES]) [S] = KM [ES] v0 = k3[Et][S] KM +[S] ([Et] –[ES]) [S] = KM[ES] k3[Et] = Vmax v0 = Vmax [S] KM +[S] ([Et] [S]) –[ES][S] = KM[ES] [ES] (KM + [S]) = [Et][S] [ES] = [Et][S] KM+ [S]
![v0 = Vmax [S] KM +[S] v0 = k3 [ES] ([Et] –[ES]) [S] = KM [ES]](http://slideplayer.it/slide/10564631/33/images/40/v0+%3D+Vmax+%5BS%5D+KM+%2B%5BS%5D+v0+%3D+k3+%5BES%5D+%28%5BEt%5D+%E2%80%93%5BES%5D%29+%5BS%5D+%3D+KM+%5BES%5D.jpg "Stato stazionario. k3. k1. k2. v0 = k3 [ES] ([Et] –[ES]) [S] = KM. [ES] v0 = k3[Et][S] KM +[S] ([Et] –[ES]) [S] = KM[ES] k3[Et] = Vmax. v0 = Vmax [S] KM +[S] ([Et] [S]) –[ES][S] = KM[ES] [ES] (KM + [S]) = [Et][S] [ES] = [Et][S] KM+ [S]")
41
v0 = Vmax [S] KM +[S] KM= k2 + k3 k1 Vmax Vmax S = Km = S 2 Km + S k3
![v0 = Vmax [S] KM +[S] KM= k2 + k3 k1 Vmax Vmax S = Km = S 2 Km + S k3](http://slideplayer.it/slide/10564631/33/images/41/v0+%3D+Vmax+%5BS%5D+KM+%2B%5BS%5D+KM%3D+k2+%2B+k3+k1+Vmax+Vmax+S+%3D+Km+%3D+S+2+Km+%2B+S+k3.jpg "v0 = Vmax [S] KM +[S] KM= k2 + k3 k1 Vmax Vmax S = Km = S 2 Km + S k3")
42
v0 = Vmax∙[S] Km +[S] Vmax = Vmax∙[S] 2 Km + [S] Km = [S]
Vmax∙(Km + [S]) = 2∙Vmax∙[S] Vmax Vmax Km + [S] = 2∙[S] Km = 2∙[S] – [S] = [S]
![v0 = Vmax∙[S] Km +[S] Vmax = Vmax∙[S] 2 Km + [S] Km = [S]](http://slideplayer.it/slide/10564631/33/images/42/v0+%3D+Vmax%E2%88%99%5BS%5D+Km+%2B%5BS%5D+Vmax+%3D+Vmax%E2%88%99%5BS%5D+2+Km+%2B+%5BS%5D+Km+%3D+%5BS%5D.jpg "Vmax∙(Km + [S]) = 2∙Vmax∙[S] Vmax Vmax. Km + [S] = 2∙[S] Km = 2∙[S] – [S] = [S]")
43
Velocità di reazione (v0) = quantità di substrato trasformato in prodotto nell’unità di tempo. Si esprime in mmoli di prodotto/L (mM)/min. Vmax = velocità massima della reazione. Saturazione dei siti attivi dell’enzima Km = costante di Michaelis-Menten È la concentrazione del substrato alla quale la velocità della reazione è metà della velocità massima. Si esprime in mmoli/L (mM)
")
44
…ancora sulla Km… k3 k1 k2 in generale, la Km rappresenta l’affinità dell’enzima per il substrato - basso valore di Km = alta affinità (è sufficiente una concentrazione bassa di substrato per avere ½ Vmax) alto valore di Km = bassa affinità (è necessaria una concentrazione alta di substrato per avere ½ Vmax) Ogni enzima ha una caratteristica Km per un determinato substrato K3 << k2 Km= k2 + k3 k1 Km= k2 k1 Km1 Km2
 alto valore di Km = bassa affinità (è necessaria una concentrazione alta di substrato per avere ½ Vmax) Ogni enzima ha una caratteristica Km per un determinato substrato. K3 << k2. Km= k2 + k3. k1. Km= k2. k1. Km1. Km2.")
45
La costante catalitica Kcat…
V0 = K3 [ES] Vmax = K3 [Et] Quando [Et] = [ES] V0 = Vmax K3 = Kcat Vmax = Kcat [Et] Kcat = Vmax [Et]

46
Kcat = Vmax [Et] Kcat = numero di turnover = numero massimo di moli di substrato convertite in prodotto nell’unità di tempo per mole di enzima (per mole di sito attivo dell’enzima) Kcat = s-1 è una misura di quanto velocemente un dato enzima può catalizzare una specifica reazione
![Kcat = Vmax [Et] Kcat = numero di turnover. = numero massimo di moli di substrato. convertite in prodotto nell’unità di tempo.](http://slideplayer.it/slide/10564631/33/images/46/Kcat+%3D+Vmax+%5BEt%5D+Kcat+%3D+numero+di+turnover.+%3D+numero+massimo+di+moli+di+substrato.+convertite+in+prodotto+nell%E2%80%99unit%C3%A0+di+tempo..jpg "per mole di enzima (per mole di sito attivo. dell’enzima) Kcat = s-1 è una misura di quanto velocemente. un dato enzima può catalizzare una specifica. reazione.")
47
5

48
Perché è utile conoscere Km, Vmax,Kcat?...
Km è unica per ogni coppia enzima-substrato

49
reazione chimica sono detti
Es: Enzimi diversi che agiscono sullo stesso substrato… Enzimi diversi che catalizzano la stessa reazione chimica sono detti ISOZIMI

50
L’equazione di Lineweaver-Burk:
v0= Vmax S KM + S Pendenza = 1 = Km V0 Vmax [S ] Vmax y = bx + c y = 1/v0 x = 1/ S b = Km/Vmax c = 1/Vmax L’equazione di Lineweaver-Burk: - è una trasformazione dell’equazione di Michaelis –Menten - permette di calcolare in modo più accurato Km e Vmax

51
Reazione catalizzata da enzimi
Effetto della TEMPERATURA

52
pH Reazione catalizzata da enzimi Effetto del stato di ionizzazione di
residui del sito attivo stato di ionizzazione delle catene laterali di tutta la catena polipeptidica: effetto sulla struttura nativa Il pH ottimale rispecchia quello dell’ambiente in cui l’enzima svolge normalmente le sue funzioni

53
Molecole che si legano all’enzima e interferiscono
Altri fattori che possono influenzare l’attività di un enzima…. Molecole che si legano all’enzima e interferiscono con la sua attività INIBITORI

54
INIBIZIONE DELL’ATTIVITA’ ENZIMATICA
DUE TIPI DI INIBIZIONE Inibizione reversibile: l’inibitore si lega e si dissocia dall’enzima rapidamente (legami non covalenti) Inibizione irreversibile: complesso stabile enzima-inibitore (legami non covalenti o covalenti) - l’enzima non recupera più la sua attività - Competitiva Non competitiva
 Inibizione irreversibile: complesso stabile enzima-inibitore (legami non covalenti o covalenti) - l’enzima non recupera più la sua attività. - Competitiva. Non competitiva.")
55
1. Inibizione competitiva: inibitore e substrato hanno lo
1. Inibizione competitiva: inibitore e substrato hanno lo stesso sito di legame Ki = costante di inibizione = costante di dissociazione del complesso EI L’inibizione competitiva può essere rimossa aumentando la concentrazione del substrato

56
Inibitori competitivi = analoghi del substrato
1. Inibizione competitiva: inibitore e substrato hanno lo stesso sito di legame Inibitori competitivi = analoghi del substrato

57
1. Inibizione competitiva: inibitore e substrato hanno lo
1. Inibizione competitiva: inibitore e substrato hanno lo stesso sito di legame Inibitore substrato enzima Effetto sulla Km KmApp = aKm a = 1 + [I] Ki

58
2. Inibizione non competitiva: l’inibitore si lega ad un sito diverso da quello del substrato, sia all’enzima libero che al complesso enzima-substrato

59
2. Inibizione non competitiva: l’enzima e il complesso ES legano l’inibitore con la stessa affinità
Effetto sulla Vmax

60
Inibitore competitivo
(I si lega solo a E) Vmax = invariata Km = aumenta Inibitore non competitivo (I si lega sia E sia ES) Vmax = diminuisce Km = invariata
 Vmax = invariata. Km = aumenta. Inibitore non competitivo. (I si lega sia E sia ES) Vmax = diminuisce. Km = invariata.")
61
INIBIZIONE ENZIMATICA IRREVERSIBILE
Diisopropil fluorofosfato L’inibitore si lega covalentemente ad un gruppo funzionale dell’enzima necessario per l’attività catalitica

62
Impiego degli inibitori irreversibili:
INIBIZIONE ENZIMATICA IRREVERSIBILE Impiego degli inibitori irreversibili: Studio dei meccanismi di reazione (identificazione di residui del sito attivo) Farmaci
 Farmaci.")
63
Gli ENZIMI ALLOSTERICI
enzimi la cui attività è influenzata da cambiamenti conformazionali mediati dall’interazione con piccole molecole (effettori o modulatori) effettori o modulatori si legano ad un sito diverso dal sito attivo effettori negativi: riducono l’attività dell’enzima effettori positivi: fanno aumentare l’attività dell’enzima sono costituiti da più subunità
 effettori o modulatori si legano ad un sito diverso dal sito attivo. effettori negativi: riducono l’attività dell’enzima. effettori positivi: fanno aumentare l’attività dell’enzima. sono costituiti da più subunità.")
64
…sito catalitico e sito del modulatore su subunità diverse…

65
ENZIMA ALLOSTERICO Velocità della reazione (v0) [S]
![ENZIMA ALLOSTERICO Velocità della reazione (v0) [S]](http://slideplayer.it/slide/10564631/33/images/65/ENZIMA+ALLOSTERICO+Velocit%C3%A0+della+reazione+%28v0%29+%5BS%5D.jpg "ENZIMA ALLOSTERICO Velocità della reazione (v0) [S]")
66
Monod-Wyman-Changeux
Modello sequenziale (Koshland) Modello concertato Monod-Wyman-Changeux (MWC)
 Modello concertato. Monod-Wyman-Changeux. (MWC)")
67
Negli enzimi allosterici la curva della velocità di reazione in funzione della concentrazione del substrato ha un andamento sigmoidale K0.5 Km

68
Effetto omotropico: il substrato stesso funge da effettore positivo
Un effettore allosterico può modificare l’ affinità dell’enzima per il substrato (K0.5) o la velocità catalitica massima (Vmax) o entrambi i parametri Effetto omotropico: il substrato stesso funge da effettore positivo Effetto eterotropico: l’effettore è un composto diverso dal substrato
 o la velocità catalitica massima (Vmax) o entrambi i parametri. Effetto omotropico: il substrato stesso funge da effettore positivo. Effetto eterotropico: l’effettore è un composto diverso dal substrato.")
69
REGOLAZIONE ENZIMATICA
MODIFICAZIONE COVALENTE: Aggiunta di un gruppo chimico alla catena laterale di specifici amminoacidi DOPO la sintesi della catena polipeptidica MODIFICAZIONI POST-TRADUZIONALI

70
H

71
Glu, Asp Arg, Lys

72
Conseguenze funzionali delle modifiche post-traduzionali:
Qual è l’effetto di una modificazione post-traduzionale?.... Conseguenze funzionali delle modifiche post-traduzionali: Attivazione/inibizione Localizzazione Acquisizione di una nuova funzione Aumento della stabilità meccanica Degradazione proteolitica …………………………..

73
La fosforilazione è un importante meccanismo di regolazione delle
vie metaboliche Enzimi che catalizzano la reazione di fosforilazione: CHINASI Enzimi che rimuovono il gruppo fosfato: FOSFATASI

74
Regolazione dell’attività della glicogeno fosforilasi
+ Pi H OPO32-

75
Attivazione degli zimogeni
REGOLAZIONE ENZIMATICA Attivazione degli zimogeni Precursore inattivo (zimogeno o proenzima) Enzima attivo Taglio proteolitico ATTIVAZIONE IRREVERSIBILE!
 Enzima attivo. Taglio proteolitico. ATTIVAZIONE IRREVERSIBILE!")
76
ATTIVAZIONE DEGLI ZIMOGENI

77
Attivazione degli zimogeni Es: attivazione del chimotripsinogeno
REGOLAZIONE ENZIMATICA Attivazione degli zimogeni Es: attivazione del chimotripsinogeno pancreas Intestino tenue

79
REGOLAZIONE ENZIMATICA
Inibitori (piccole molecole che interferiscono con la catalisi- inibizione reversibile/irreversibile) Modulatori allosterici (effettori positivi/negativi-cooperatività- effetti omotropici/eterotropici) Modificazioni covalenti (modificazioni post-traduzionali reversibili. Molti enzimi del metabolismo regolati per fosforilazione/defosforilazione) Attivazione di zimogeni (conversione di un precursore inattivo in un enzima attivo mediante taglio proteolitico) Isozimi Regolazione dell’espressione genica (quantità diverse di enzima prodotte in risposta a stimoli specifici) -Regolazione del turnover (velocità di sintesi e degradazione) Compartimentazione (-enzimi coinvolti nello stesso processo formano complessi multiproteici -enzimi coinvolti in vie metaboliche diverse operano in compartimenti diversi della cellula)
 Modulatori allosterici. (effettori positivi/negativi-cooperatività- effetti omotropici/eterotropici) Modificazioni covalenti. (modificazioni post-traduzionali reversibili. Molti enzimi del metabolismo. regolati per fosforilazione/defosforilazione) Attivazione di zimogeni. (conversione di un precursore inattivo in un enzima attivo mediante taglio. proteolitico) Isozimi. Regolazione dell’espressione genica. (quantità diverse di enzima prodotte in risposta a stimoli specifici) -Regolazione del turnover. (velocità di sintesi e degradazione) Compartimentazione. (-enzimi coinvolti nello stesso processo formano complessi multiproteici. -enzimi coinvolti in vie metaboliche diverse operano in compartimenti. diversi della cellula)")
Presentazioni simili










>")








