Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
In molti casi un grafico di Scatchard può essere curvo piuttosto che lineare. Questo succede in due casi classici: 1 - Classi multiple di siti indipendenti 2 - Interazioni tra siti di legame, in particolare a - Legami cooperativi b - Legami anticooperativi GRAFICI DI SCATCHARD CURVI

2
La figura illustra un grafico bifasico nel caso di due classi di siti indipendenti: l’intercetta con l’asse delle ordinate è uguale a CLASSI MULTIPLEDI SITI INDIPENDENTI

3
In prima approssimazione possiamo assumere che questo termine è dominato dal più basso valore di k (definito come k 1 ) e stimare che l’intercetta sia uguale a Allo stesso modo l’intercetta sull’asse delle ascisse della tangente alla curva nel tratto per [L] che tende a 0 (cioè molto piccolo) sarà la prima stima di n 1. Con i valori stimati di n 1 e k 1 possiamo sottrarre dai dati il contributo del legame più forte (valore più piccolo di k) e possiamo costruire un nuovo grafico che può essere analizzato con l’espressione lineare in modo da stimare i valori di n 2 e k 2
![In prima approssimazione possiamo assumere che questo termine è dominato dal più basso valore di k (definito come k 1 ) e stimare che l’intercetta sia uguale a Allo stesso modo l’intercetta sull’asse delle ascisse della tangente alla curva nel tratto per [L] che tende a 0 (cioè molto piccolo) sarà la prima stima di n 1.](http://images.slideplayer.it/39/10920328/slides/slide_3.jpg "Con i valori stimati di n 1 e k 1 possiamo sottrarre dai dati il contributo del legame più forte (valore più piccolo di k) e possiamo costruire un nuovo grafico che può essere analizzato con l’espressione lineare in modo da stimare i valori di n 2 e k 2.")
4
il processo può essere iterato per ottenere una nuova stima di n 2 e k 2. Durante tutto il calcolo si deve usare il vincolo che n 1 + n 2 sia uguale al valore dell’intercetta con l’asse delle ascisse ed il processo terminerà quando il valore di trovato sarà uguale all’intercetta sull’asse delle ordinate.

5
La fig. mostra i dati per il legame di Mn +2 ad una porzione di t-RNA in trietanolammina 0.1 M. Questo pezzo di RNA contiene sia regioni a singola elica che regioni a doppia elica. Sono state trovate due classi di siti n 1 =6 ed n 2 = 10; le costanti di dissociazione sono k 1 = 14 M e k 2 = 200 M.

6
i parametri ottenuti dall’analisi di un grafico di Scatchard multifasico sono gli unici possibili? Una risposta può essere data andando a vedere come cambiano i valori delle costanti k i al variare dei valori del numero di siti di legame per ogni classe di siti. Se la variazione è piccola possiamo ritenere che i dati sono stati interpretati correttamente. P.es., nel caso precedente una variazione di 1 nella variabile n 1 dà una variazione minore del 50% nei valori di k 1.

7
Un’altra domanda che ci si può porre è se i dati possano essere descritti altrettanto bene supponendo l’esistenza di più di due classi di siti. Naturalmente più alto è il numero di parametri disponibili per interpolare dei dati con un modello, migliore sarà l’accordo tra teoria ed esperimenti. Il procedimento migliore è quello di trattare i dati, all’interno dei limiti dell’errore sperimentale, con il minor numero possibile di assunzioni e di parametri. Questo modo di procedere dà un quadro circa il minor numero possibile di caratteristiche del sistema che forse sono anche quelle dominanti.

8
INTERAZIONI TRA SITI E’ possibile che il legame di un ligando alteri l’affinità della macromolecola per il successivo ligando e così via producendo una variazione continua del valore delle costanti di dissociazione microscopiche. Nel caso più semplice di una sola classe di siti identici e dipendenti possiamo definire k o la costante di dissociazione microscopica per = 0. Quando aumenta, l’interazione tra i siti fa variare il valore di k.

9
Se G° è la variazione di energia libera standard per la dissociazione di un ligando legato: G° = G o ° + RT ( ) dove G o ° = - RTlnk o e ( ) è una funzione che, per definizione, prende in considerazione gli effetti delle interazioni tra siti che variano col grado di saturazione. Allora: dove ( ) = 0 per = 0 In modo che:
 = 0 per = 0 In modo che:.")
10
Se ( ) è una funzione decrescente di allora k( ) cresce verso la saturazione: in questo caso il grafico di Scatchard sarà curvo con la concavità verso l’alto. D’altra parte se ( ) aumenta all’aumentare di allora il grafico di Scatchard avrà un massimo con la concavità verso il basso.
 aumenta all’aumentare di allora il grafico di Scatchard avrà un massimo con la concavità verso il basso..")
11
EFFETTI STATISTICI ED ENERGIA DI INTERAZIONE. consideriamo una macromolecola che lega quattro ligandi L: se tutti i siti sono identici ed indipendenti e legano L con una costante microscopica k, allora attraverso il trattamento statistico visto precedentemente è possibile conoscere le quattro costanti macroscopiche: K 1 <K 2 <K 3 <K 4 ed il processo, dal punto di vista delle costanti macroscopiche, sembra essere sempre più debole al crescere della saturazione anche se la costante microscopica è sempre la stessa: dal punto di vista delle costanti macroscopiche gli effetti statistici introducono una “anticooperatività apparente”.

12
In un sistema veramente cooperativo, una volta effettuata la correzione per gli effetti statistici, la costante di dissociazione di uno o più stadi successivi decresce al crescere della saturazione. Nell'esempio della macromolecola con 4 siti questo vuol dire che, se c'è cooperatività tra il primo ed il secondo stadio, si ha: Se tutti gli stadi coinvolgono un legame via via più forte allora:

13
La forza della cooperatività coinvolta nel legare due ligandi può essere espressa in unità di energia libera con una procedura semplice. Supponiamo che: G° i = RT ln K i sia la variazione di energia libera standard apparente nell'equilibrio di legame dell'i-esimo ligando della serie. (Si tenga presente che K i è una costante di dissociazione cosicchè -RT ln K i è la variazione di energia libera associata con il processo di dissociazione; quindi +RT ln K i si riferisce al processo di associazione). Tale variazione contiene una componente puramente statistica data da:
. Tale variazione contiene una componente puramente statistica data da:.")
14
Per tener conto di questo termine definiamo una variazione di energia libera standard intrinseca relativa al legame dell'i-esimo ligando, G'°: Definiamo allora l'energia di interazione, G I,ij, per sito, come la differenza tra l'energia libera intrinseca di associazione dell'i-esimo e quella del j-esimo ligando. Questa energia di interazione sarà data da:

15
Con questa definizione di G I,ij, se il j-esimo ligando lega più fortemente dell’i-esimo (j>i) allora in un sistema cooperativo si ha G I,ij < 0. Si noti pure che se i due siti hanno la stessa costante di dissociazione intrinseca allora i due termini logaritmici dell'espressione precedente si cancellano e G I,ij = 0. Nel caso dell'ossigeno legato all'emoglobina umana, l'equazione precedente fornisce un valore di G I,ij - 2 kcal/mol per sito per i = 1 e j = 4. Questo significa che le interazioni sito-sito stabilizzano una molecola di ossigeno legata in un tetramero di emoglobina satura di circa 2 kcal/mol rispetto ad una molecola di ossigeno legato ad una emoglobina che ha tre siti liberi.

16
INTERAZIONE DI LIGANDI GRANDI CON CATENE DI TIPO RETICOLO LINEARE. Un caso di notevole interesse è dato dall'associazione di “ligandi grandi” con catene di tipo "reticolo lineare" come p.es. la doppia elica degli acidi nucleici. Nel caso del DNA, I "grandi ligandi" includono le poliammine, gli istoni, alcune proteine capaci di interagire con l'elica del DNA. Questi sistemi hanno delle proprietà caratteristiche che sono abbastanza diverse da quelle viste finora.

17
Modello di interazione tra la doppia elica del DNA ed una poliammina

18
IL "RETICOLO LINEARE": CARATTERISTICHE DI TIPO STATISTICO Consideriamo un reticolo omogeneo costituito da N unità ripetitive identiche. Ad esempio, in un'elica di DNA le unità ripetitive potrebbero essere i gruppi carichi fosfato o i residui deossiribosici. Si assuma che un ligando L occupi l unità consecutive del reticolo.

19
In un reticolo vuoto ci saranno N- l +1 siti potenziali che la prima molecola di ligando può occupare. In questo modo all'inizio della titolazione ci sono molti più siti potenziali dei N/ l siti che possono essere occupati a saturazione.

20
La figura illustra la complessità statistica del reticolo per il caso N = 12 e l = 3. Siano le moli di L legato per mole di reticolo; a saturazione si avrà = N/l = 4. All'inizio della titolazione tuttavia ci sono N-l+1 (10) siti potenziali per reticolo.
 siti potenziali per reticolo..")
21
Per calcolare il comportamento del legame di ligandi su reticolo partiamo dalla definizione di costante microscopica: dove [sito] f rappresenta la concentrazione dei siti liberi, [sito] b rappresenta la concentrazione dei siti occupati ed [L] è la concentrazione del ligando libero.
![Per calcolare il comportamento del legame di ligandi su reticolo partiamo dalla definizione di costante microscopica: dove [sito] f rappresenta la concentrazione dei siti liberi, [sito] b rappresenta la concentrazione dei siti occupati ed [L] è la concentrazione del ligando libero.](http://images.slideplayer.it/39/10920328/slides/slide_21.jpg "Per calcolare il comportamento del legame di ligandi su reticolo partiamo dalla definizione di costante microscopica: dove [sito] f rappresenta la concentrazione dei siti liberi, [sito] b rappresenta la concentrazione dei siti occupati ed [L] è la concentrazione del ligando libero.")
22
La concentrazione dei siti liberi è espressa dal prodotto della concentrazione totale della macromolecola, che rappresenta il reticolo lineare, per il numero medio per macromolecola di siti liberi che abbiano una lunghezza l : Poichè

23
Il nostro scopo è quello di calcolare un espressione per. Faremo questo calcolando dapprima la probabilità P l che, partendo da una qualsiasi posizione del reticolo, l unità consecutive del reticolo non siano occupate da un ligando. Questa probabilità moltiplicata per N dà il numero medio di siti liberi e lunghi l per mole di reticolo. La probabilità P l può essere scritta come: dove P f è la probabilità che una unità del reticolo scelta a caso non sia occupata e P ff è la probabilità condizionale che una certa unità libera sia seguita da un'altra unità libera.

24
Poiché la frazione di unità di reticolo occupate è: la frazione di unità libere è: quindi la probabilità di trovare una unità occupata è: mentre

25
La probabilità condizionale P ff è anche la frazione del totale delle unità libere che sono precedute da altre unità libere. Questa è data dal numero totale di unità di reticolo libere meno il numero di unità di reticolo libere che sono precedute da unità occupate, il tutto diviso per il numero totale di unità libere: dove P bf è la probabilità condizionale che una unità di reticolo libera segua l’ultimo segmento ( l -esimo) di un ligando legato e P b / l è la probabilità che un’unità di reticolo legata sia occupata dall’ultimo ( l - esimo) segmento del ligando.
 di un ligando legato e P b / l è la probabilità che un’unità di reticolo legata sia occupata dall’ultimo ( l - esimo) segmento del ligando..")
26
Poiché non vi sono interazioni tra ligandi legati è chiaro che P ff = P bf ; cioè la probabilità che un ligando si disponga dopo un sito libero o dopo un sito occupato dalla coda di un altro ligando è la stessa. Quindi: da cui

27
Da cui: Possiamo ora scrivere l’espressione di

28
Per trovare questo risultato abbiamo ignorato gli effetti terminali sicchè a rigore l’espressione precedente vale solo per un reticolo infinitamente lungo. Quando l = 1 l’equazione si riduce a (n = N ):
:.")
29
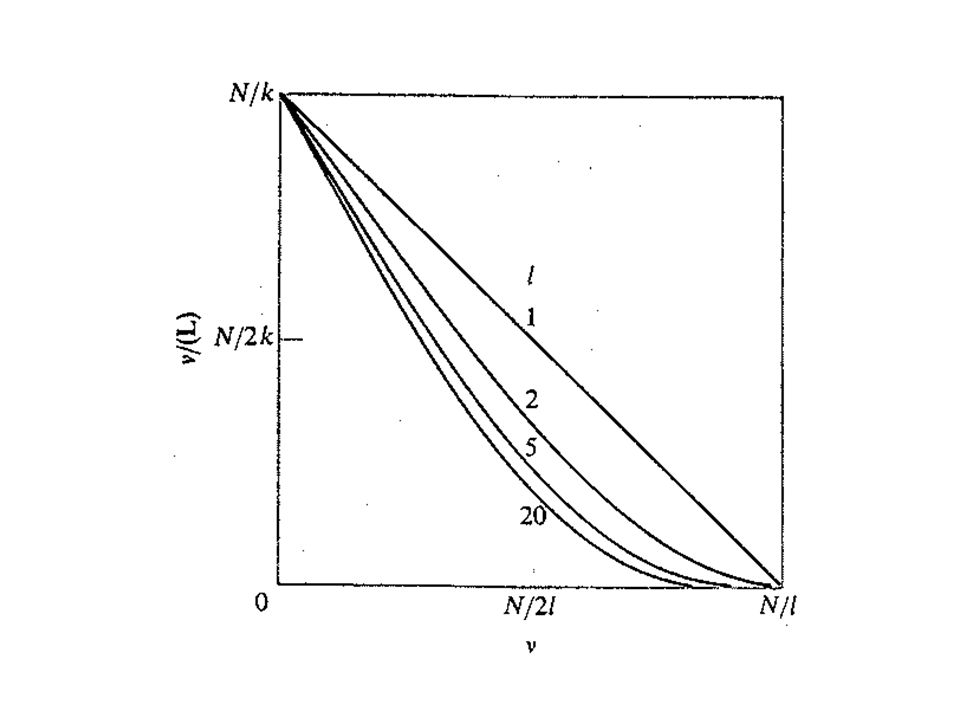
quando l > 1, l’ultimo termine dell’espressione è sempre minore dell’unità e varia al variare di. Questo dà luogo ad una marcata non-linearità nei grafici di Scatchard ed inoltre le curve si trovano al di sotto del caso l = 1. La figura mostra alcuni grafici di Scatchard calcolati per valori di l tra 1 e 20 con k = 1 M. Per facilitare il confronto tra i grafici l’ascissa è data in unità di 1/ l.

31
Al crescere di l la concavità verso l’alto delle curve è più marcata perché il reticolo resiste, dal punto di vista entropico, alla saturazione e questa resistenza diviene più pronunciata al crescere di l. Il gran numero di microstati che si generano ad una certa saturazione parziale dà un forte contributo entropico all’energia libera. Questo contributo viene naturalmente perso non appena il legame procede verso la saturazione del reticolo, che è costituita da un solo microstato. Ne consegue che in pratica per ligandi molto grandi la saturazione non è un processo possibile; p.es. passando da l = 10 ad l = 20 la concentrazione di ligando deve variare di 10 e 100 volte rispettivamente per passare da una saturazione parziale dell’80% ad una del 90%.

32
L’intercetta delle ascisse del grafico di Scatchard è N/ l che corrisponde a n nell’espressione per l = 1. Però l’intercetta sulle ordinate è N/k che non è la stessa cosa di n/k. Questa distinzione è importante quando si ottengono parametri di legame da un grafico di Scatchard che coinvolge interazioni con una catena di tipo “reticolo lineare omogeneo”.

33
La freccia denota il punto di saturazione completa. Le curve tratteggiate denotano la sensibilità dei dati a variazioni di l tenendo costante k. In questo caso l = 7.8 e k = 7.1 M. La fig. seguente mostra un caso reale di substrato che lega un reticolo lineare: l'esapeptide -DNP-Lys-(lys) 5 che interagisce con l'elica di poli(rI):poli(rC).
 5 che interagisce con l elica di poli(rI):poli(rC)..")
Presentazioni simili


















>")
