Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
dopo la percezione…

2
Risposte tra 1 min e 2h The perception step is followed by activation of protein kinases (PK) or inhibition of protein phosphatases (PP) that, in turn, trigger the Ca2+ influx. Within the first 5 min, nearly 20 phosphoproteins showed an increase in phosphorylation status Extracellular Ca2+ influx, depending on PK activation, triggers ROS production, MAPK activation, PK activation upstream of the NO production, anion effluxes and plasma membrane depolarization, glucose (Glc) import inhibition, microtubule depolymerization, and may lead to the H+-ATPase inhibition
 or inhibition of protein phosphatases (PP) that, in turn, trigger the Ca2+ influx. Within the first 5 min, nearly 20 phosphoproteins showed an increase in phosphorylation status Extracellular Ca2+ influx, depending on PK activation, triggers ROS production, MAPK activation, PK activation upstream of the NO production, anion effluxes and plasma membrane depolarization, glucose (Glc) import inhibition, microtubule depolymerization, and may lead to the H+-ATPase inhibition")
3
La risposta locale

5
I ROS La famiglia dei ROS intesa in senso stretto consiste in:
Anione superossido O2.- Idroperossil radicale HO2. Perossido d'idrogeno H2O2 Idrossil radicale HO. Il superossido, la sua forma protonata e l'idrossil radicale hanno un breve tempo di vita mentre l' H2O2 è relativamente stabile e può passare attraverso le membrane Tra i ROS, l'idrossil-radicale è particolarmente tossico a causa della sua straordinaria capacità di reagire spontaneamente con diverse molecole organiche come i fenoli, gli acidi grassi, le proteine e gli acidi nucleici

6
Pathways for oxygen and nitrogen free radical species

8
ROS nell'interazione pianta-patogeno
I ROS sono fattori chiave nella predisposizione e nel mantenimento sia dell'inaccessibilità che dell'accessibilità dell'ospite per i patogeni Infatti da un lato i ROS sono implicati nella signal transduction e nell'esecuzione delle reazioni di difesa come nel rafforzamento della parete cellulare e nella rapida morte cellulare (HR). Dall'altro i ROS si accumulano anche nelle reazioni compatibili in cui sembrano paradossalmente connessi con la limitazione della diffusione della morte cellulare Diversi tipi di ROS possono stimolare effetti diametralmente opposti in dipendenza sia della loro distribuzione spaziotemporale che delle loro concentrazioni subcellulari
. Dall altro i ROS si accumulano anche nelle reazioni compatibili in cui sembrano paradossalmente connessi con la limitazione della diffusione della morte cellulare. Diversi tipi di ROS possono stimolare effetti diametralmente opposti in dipendenza sia della loro distribuzione spaziotemporale che delle loro concentrazioni subcellulari.")
9
ROS nell'interazione pianta-patogeno
I ROS svolgono diversi ruoli nell'interazione che vanno da un effetto benefico che porta allo stabilirsi di una reazione incompatibile ad un effetto coadiuvante l'infezione (patogeni necrotrofi) Le cellule vegetali rispondono alle infezioni batteriche con un rapido e transiente accumulo bifasico dei ROS Mentre il primo ciclo di accumulo avviene sia nelle interazioni compatibili che in quelle incompatibili la seconda, prolungata, fase precede la morte cellulare e dipende dalla presenza di un'interazione specifica R/Avr che porta all'incompatibilità
 Le cellule vegetali rispondono alle infezioni batteriche con un rapido e transiente accumulo bifasico dei ROS. Mentre il primo ciclo di accumulo avviene sia nelle interazioni compatibili che in quelle incompatibili la seconda, prolungata, fase precede la morte cellulare e dipende dalla presenza di un interazione specifica R/Avr che porta all incompatibilità.")
10
Il burst ossidativo Il burst ossidativo è una risposta comune a tutti i tipi di stress sia biotico che abiotico ed includono l’HR e la risposta all’ozono Ad esempio in tabacco e Arabidopsis un burst ossidativo bifasico o un prolungato accumulo di ROS risultano nell’attivazione della PCD In diverse specie vegetali è stata osservata una chiara correlazione spaziale tra l’accumulo di ROS e le lesione cellulari In tabacco, betulla e pomodoro ad es. c’è una chiara correlazione tra il perossido d’idrogeno e le lesioni tardive mentre l’aso è maggiormente collegato alla morte cellulare in Arabidopsis e malva, indicando un ruolo specie-specifico dei ROS nella modulazione della PCD

11
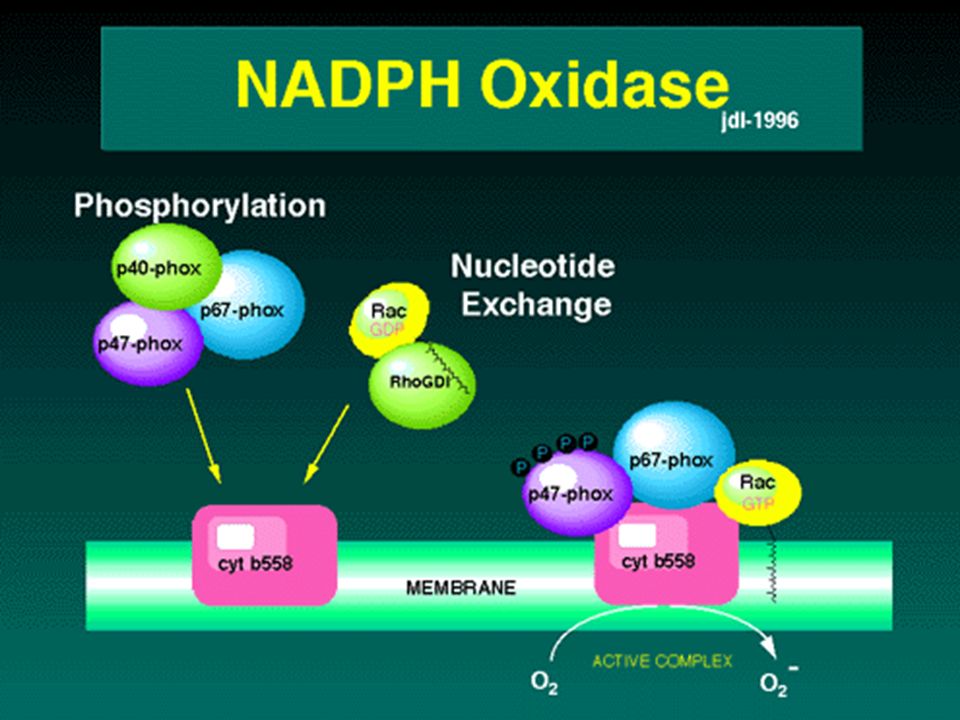
NADPHox come sorgente di ROS
L'accumulo molto precoce di H2O2 nell'apoplasto dopo il contatto col patogeno suggerisce che almeno in parte questo derivi dalla dismutazione dell'anione superossido prodotto dal complesso della NADPH ox Infatti in tabacco l'esposizione al patogeno causa una up-regolazione di due isoforme di NADPH ox Quindi la produzione di aso e la sua dismutazione regolano la presenza di ROS nell'apoplasto, quando è molto attiva la NADPHox e il tasso di dismutazione è alto si accumula H2O2 mentre quando questo tasso è basso si accumula aso

13
La regolazione della NADPHox
La precoce induzione di rapidi flussi ionici attraverso la membrana plasmatica sono coinvolti direttamente nell'induzione del burst ossidativo Le proteine RBOH contengono dei Ca2+ binding domain e non hanno le subunità citoplasmatiche separate come quelle delle cellule animali (p67, p47...) quindi gli ioni calcio svolgono un ruolo importante nel controllo della produzione di aso Anche gli spostamenti anionici (Cl-; NO3-) sono coinvolti nell'attivazione del burst ox Altri importanti regolatori del complesso NADPHox sia nelle piante che negli animali sono le protein kinasi e le fosfatasi, le small GTPasi e le fosfolipasi. Infatti una small GTPasi della famiglia Rac è implicata nella produzione di ROS e nella PCD NADPH provided by the pentose phosphate pathway induced in the first minutes of plant-pathogen interaction
 quindi gli ioni calcio svolgono un ruolo importante nel controllo della produzione di aso. Anche gli spostamenti anionici (Cl-; NO3-) sono coinvolti nell attivazione del burst ox. Altri importanti regolatori del complesso NADPHox sia nelle piante che negli animali sono le protein kinasi e le fosfatasi, le small GTPasi e le fosfolipasi. Infatti una small GTPasi della famiglia Rac è implicata nella produzione di ROS e nella PCD. NADPH provided by the pentose phosphate pathway induced in the first minutes of plant-pathogen interaction.")
14
..dopo l’attivazione Ci sono almeno 3 vie che indipendentemente portano ad una riprogrammazione trascrizionale in seguito all’attivazione della difesa La prima è legata ai geni EDS1 (enhanced disease susceptibility) o PAD4 che interagiscono fisicamente tra loro in vivo EDS1/PAD4 interagiscono principalmente con le proteine CC-NB- LRR La riprogrammazione trascrizionale seguente al riconoscimento del patogeno è dipendente dall’accumulo a livello nucleare di EDS1
 o PAD4 che interagiscono fisicamente tra loro in vivo. EDS1/PAD4 interagiscono principalmente con le proteine CC-NB- LRR. La riprogrammazione trascrizionale seguente al riconoscimento del patogeno è dipendente dall’accumulo a livello nucleare di EDS1.")
15
NDR1 La seconda è legata a NDR1 (non race specific disease resistance) che codifica per una glicosilfasfatidilinositolo(GPI) anchored protein Negli animali queste proteine si trovano nei rafts lipidici (specialised membrane domains enriched in certain lipids cholesterol and proteins ) e sono associate con complessi recettoriali. NDR1 interagisce principalmente con le proteine TIR-NB-LRR (ad es. interagisce con RIN4 e con RPM1)
 e sono associate con complessi recettoriali. NDR1 interagisce principalmente con le proteine TIR-NB-LRR (ad es. interagisce con RIN4 e con RPM1)")
16
RPW8 Una terza via è rappresentata dai geni R RPW8 1 e 2 che conferiscono resistenza a numerose powdery mildews (oidio) e codificano per delle proteine con domini CC ma senza le regioni NB e LRR
 e codificano per delle proteine con domini CC ma senza le regioni NB e LRR.")
17
Ricapitolando…. Dopo la percezione dei PAMP del patogeno o dopo un interazione avr/R si depolarizza la membrana e si ha un efflusso di ioni Ca con attivazione di calmoduline/calmoduline kinasi e canali ionici di membrana Questi fattori contribuiscono all’attivazione della NADPHox (burst ossidativo) e di altre cascate MAP kinasiche nonché all’attivazione della NO sintasi Queste pathway contribuiscono indipendentemente o sinergicamente all’attivazione dei fattori EDS1/PAD4 o NDR1 Cosa controllano questi fattori? In primis la sintesi di fitormoni
 e di altre cascate MAP kinasiche nonché all’attivazione della NO sintasi. Queste pathway contribuiscono indipendentemente o sinergicamente all’attivazione dei fattori EDS1/PAD4 o NDR1. Cosa controllano questi fattori In primis la sintesi di fitormoni.")
18
Il ruolo degli ormoni nell’interazione pianta-patogeno
Il complesso interplay degli jasmonati con l’acido salicilico e l’etilene fornisce alle piante un potenziale regolativo in grado di controllare il destino dell’interazione pianta-patogeno Gli ormoni acido jasmonico, salicilico e l’etilene sono i principali regolatori dell’immunità innata nelle piante

19
Acido Jasmonico

20
Ruolo del JA A seconda del tipo di interazione ospite-patogeno il JA, l’SA e l’Et sono coinvolti in modo differenziale nella PTI, infatti le pathway di signalling difensivo che si attivano sono diverse a seconda dello “stile di vita” del patogeno Le risposte difensive stimolate dal SA sono normalmente associate con la PCD e questo tipo di risposta è effettiva contro i biotrofi ma è non efficiente o addirittura peggiorativa nel caso dei necrotrofi

21
Il ruolo del JA Le risposte difensive stimolate dal JA sono invece effettive contro i necrotrofi, infatti mutanti jar1 (ridotta sensibilità al metiljasmonato) e fad3-8 (ridotta biosintesi di JA) di At risultano essere suscettibili ad alcune specie normalmente non patogeniche di Pythium, un oomicete necrotrofico Il mutante JA-insensitive coi1 mostra aumentata suscettibilità a Erwinia carotovora ed ai funghi nectrotrofi Alternaria brassicola e Botrytis cinerea Un overespressione di una JA carbossimetil-transferasi aumenta i livelli endogeni di MeJA e risulta in una maggiore resistenza a B. cinerea
 e fad3-8 (ridotta biosintesi di JA) di At risultano essere suscettibili ad alcune specie normalmente non patogeniche di Pythium, un oomicete necrotrofico. Il mutante JA-insensitive coi1 mostra aumentata suscettibilità a Erwinia carotovora ed ai funghi nectrotrofi Alternaria brassicola e Botrytis cinerea. Un overespressione di una JA carbossimetil-transferasi aumenta i livelli endogeni di MeJA e risulta in una maggiore resistenza a B. cinerea.")
22
Jasmonate signaling La catena di trasduzione del segnale che lega la sintesi del JA ai cambiamenti da questo indotti nell’espressione genica consiste in un quartetto di fattori interagenti: Un JA signal SCF-COI1 – E3 ubiquitina ligasi SCF complex JAZ proteins – proteine repressore targeted da COI1 per la degradazione attraverso il proteasome 26S Fattori di trascrizione (TF) che regolano positivamente l’espressione dei JA responsive genes
 che regolano positivamente l’espressione dei JA responsive genes.")
23
Jasmonate signaling Le cellule che contengono bassi livelli di JA i geni responsive al JA sono repressi (OFF) dalle JAZ proteins che bloccano l’attività dei fattori di trascrizione (TF: e.g. MYC2) coinvolto nell’espressione degli early response gene La transizione dallo stato represso a quello attivo (ON) è iniziata da stimoli ambientali (e.g. biotic stress) che inducono un aumento di forme bioattive del JA
 dalle JAZ proteins che bloccano l’attività dei fattori di trascrizione (TF: e.g. MYC2) coinvolto nell’espressione degli early response gene. La transizione dallo stato represso a quello attivo (ON) è iniziata da stimoli ambientali (e.g. biotic stress) che inducono un aumento di forme bioattive del JA.")
24
Jasmonate signaling I JA bioattivi (cerchio arancio) come il coniugato JA-Ile si legano al dominio LRR di COI1. L’interazione delle JAZ con il COI1 legato al JA porta alla formazione di un complesso ternario JA-JAZ-COI1 in cui JAZ viene poliubiquinata (cerchi neri) e seguentemente degradata dal proteasoma 26S
 e seguentemente degradata dal proteasoma 26S.")
25
Jasmonate signaling Il signaling è attenuato dal metabolismo dei JA bioattivi verso le forme inattive dell’ormone (esagono arancio) cosi come dalla sintesi de novo delle proteine JAZ Alcuni batteri fitopatogeni come P. syringae producono la coronatina un analogo strutturale del JA-Ile Il legame ad alta affinità della COR con il complesso COI1- JAZ promuove la distruzione proteolitica di JAZ
 cosi come dalla sintesi de novo delle proteine JAZ. Alcuni batteri fitopatogeni come P. syringae producono la coronatina un analogo strutturale del JA-Ile. Il legame ad alta affinità della COR con il complesso COI1- JAZ promuove la distruzione proteolitica di JAZ.")
26
Modulazione del sistema a 4
Lo switch tra lo stato bloccato e quello attivo dell’espressione dei geni JA responsive è quindi stimolato dalla proteolisi di JAZ. Questa transizione è iniziata in risposta a diversi tipi di stress ed è dovuta JA stimola il turnover di jaz attraverso un meccanismo che richiede COI1 e il proteasoma 26S JA promuove una interazione diretta tra COI1 e JAZ Questo semplice sistema di switch ormonale fornisce uno stretto accoppiamento temporale tra l’aumento intracellulare di JA e i cambiamenti nella gene expression. Ad es. un rapido accumulo di JA (>5 min) in seguito a wounding avviene praticamente in coincidenza con l’espressione degli early JA-responsive genes
 in seguito a wounding avviene praticamente in coincidenza con l’espressione degli early JA-responsive genes.")
27
I JAs bioattivi Il neo sintetizzato JA è soggetto a diverse modifiche enzimatiche che danno origine a una pletora di derivati tra cui quelli con AA come l’Ile o la valina. Nonostante molti JA non coniugati siano attivi per se come il JA, il MeJA e le OPDA non sono attivi nell’interazione COI1-JAZ Infatti il segnale primario per questo complesso sono i coniugati JA-Ile o JA-Val mentre i non coniugati potrebbero agire attraverso un sistema diverso da quello COI1-JAZ I mutanti jar1 sono difettivi nella coniugazione di JA con Ile, infatti JAR1 è una GH3 conjugating enzyme che esibisce una elevata affinità di substrato sia con JA che con Ile I fenotipi associati con i mutanti jar1 indicano che JA-Ile sono fondamentali per la resistenza ai necrotrofi e a diversi tipi di stress abiotico

28
I JAs bioattivi La coronatina sopprime le risposte immunitarie dell’ospite attivando la JA signaling pathway attraverso un COI1- dependent manner. La similarità strutturale di COR con JA-Ile suggerisce che la tossina agisca mimando JA-Ile Infatti COR è capace di stimolare l’interazione COI1-JAZ e inoltre è 100 volte più attiva di JA-Ile nel farlo La degradazione di JAZ mediata da COR fornisce un classico esempio di come i patogeni possano alterare le pathway di signaling ormonale (dell’acido salicilico) nell’ospite per promuovere l’infezione
 nell’ospite per promuovere l’infezione.")
29
L’acido salicilico EDS1/PAD4 è necessario per due passaggi. È richiesto per l’attivazione di EDS5 EDS5 agisce a monte di sid2 nel regolare la biosintesi di SA, infatti la sua espressione è inalterata nel mutante sid2 (sid codifica per una isocorismato sintasi) SA può attivare l'espressione delle PR proteins e la resistenza attraverso 2 meccanismi. Il primo richiede i geni NPR1 e TAG2 che contrastano l'effetto inibitorio del soppressore inducibile di NPR1 (SNI1) Un segnale lipidico generato da SFD-1 è necessario per l'attivazione della pathway del SA dip. da NPR1
 SA può attivare l espressione delle PR proteins e la resistenza attraverso 2 meccanismi. Il primo richiede i geni NPR1 e TAG2 che contrastano l effetto inibitorio del soppressore inducibile di NPR1 (SNI1) Un segnale lipidico generato da SFD-1 è necessario per l attivazione della pathway del SA dip. da NPR1.")
30
Biosintesi del SA L'attivazione della pathway di signalling mediata dai geni R è capace di attivare la sintesi di SA SA si accumula sia a livello locale che sistemico in seguito a un infezione e induce la sintesi di diverse PR proteins La mancata produzione di SA previene l'induzione della SAR Viene degradato dalla salicilato idrossilasi codificata dal gene nahG presente anche in molti batteri fitopatogeni

31
L’acido salicilico EDS1/PAD4 è necessario per due passaggi. È richiesto per l’attivazione di EDS5 EDS5 agisce a monte di sid2 nel regolare la biosintesi di SA, infatti la sua espressione è inalterata nel mutante sid2 (sid codifica per una isocorismato sintasi) SA può attivare l'espressione delle PR proteins e la resistenza attraverso 2 meccanismi. Il primo richiede i geni NPR1 e TAG2 che contrastano l'effetto inibitorio del soppressore inducibile di NPR1 (SNI1) Un segnale lipidico generato da SFD-1 è necessario per l'attivazione della pathway del SA dip. da NPR1
 SA può attivare l espressione delle PR proteins e la resistenza attraverso 2 meccanismi. Il primo richiede i geni NPR1 e TAG2 che contrastano l effetto inibitorio del soppressore inducibile di NPR1 (SNI1) Un segnale lipidico generato da SFD-1 è necessario per l attivazione della pathway del SA dip. da NPR1.")
32
NPR1: non-expresser of PR genes
Anche noto come NIM1 o SAI1 Regolatore Positivo della SAR Agisce a valle di SA, a monte dei PR genes npr1 mutanti sono suscettibili a diversi patogeni Over-espressione di NPR1 genera broad-spectrum resistance Unico, ma simile a NFκ-B (regulator of immunity in animals) Avr R gene SA NPR1 PR-1 PR-2 PR-5 SAR
 Avr. R gene. SA. NPR1. PR-1 PR-2 PR-5. SAR.")
33
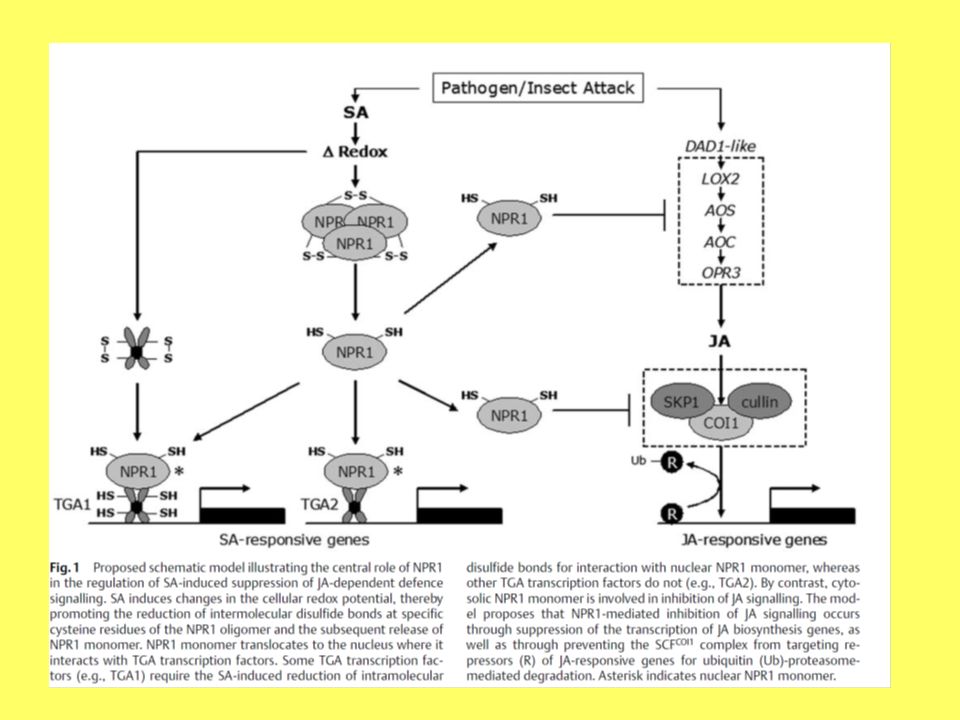
Relazione tra NPR1 e i ROS
Nelle piante lo stato redox è capace di regolare NPR1, infatti questo normalmente si accumula come oligomero che si mantiene inattivo grazie a dei ponti disolfuro nel citosol Dopo l'elicitazione la riduzione dei ponti disolfuro rilascia delle unità monomeriche che traslocano nel nucleo e interagiscono con la forma ridotta del fattore di trascrizione TGA1 che a sua volta attiva l'espressione dei geni di difesa dipendenti da SA

34
I fattori TGA Interagiscono con NPR1 Sono bZIP transcription factors
6 membri in Arabidopsis (TGA1- 6) Si legano ad elementi as-1
 Si legano ad elementi as-1.")
35
NPR1 non solo induce i PR genes ma anche i secretion-related genes
Questi secretion-related genes sono necessari per un trasporto efficiente delle PR proteins e per l'induzione della SAR NPR1 interagisce con un cis-elemento (TL1) per indurre I secretion-related genes separatamente dai PR gene element (as- 1)
 per indurre I secretion-related genes separatamente dai PR gene element (as- 1)")
36
Il ruolo dell’etilene nel interazione ospite-patogeno
Il fitormone etilene è il principale modulatore in molti aspetti della vita di una pianta tra cui i vari meccanismi attraverso i quali una pianta reagisce all’attacco di un patogeno Il signaling intracellulare “acceso” dall’Et porta ad una cascata di fattori di trascrizione che consiste primariamente dei regolatori EIN3 e secondariamente dei fattori ERF Questi controllano l’espressione di diversi geni coinvolti in vari aspetti della resistenza sistemica indotta Sono presenti livelli significativi di cross-talk tra le diverse pathways di risposte difensive controllate dal SA e dal JA che alla fine risulta in una risposta di difesa differenziata

37
La biosintesi di etilene
L’Et è sintetizzato a partire dalla metionina che è convertita ad S- adenosilmetionina (AdoMet) dalla S-adomet sintasi (ADS). Adomet è il principale donatore di gruppi metilici nelle piante ed è coinvolto nelle reazioni di metilazione dei lipidi, proteine o acidi nucleici L’adomet è convertita dall’enzima ACS (ACC sintasi) a metiltioadenosina che attraverso il ciclo di Yang è trasformata in aminociclopropano-1-carbossilato (ACC) il precursore dell’Et L’ACC è ossidato dalla ACC ox (ACO) per formare Et, cianuro e CO2
 dalla S-adomet sintasi (ADS). Adomet è il principale donatore di gruppi metilici nelle piante ed è coinvolto nelle reazioni di metilazione dei lipidi, proteine o acidi nucleici. L’adomet è convertita dall’enzima ACS (ACC sintasi) a metiltioadenosina che attraverso il ciclo di Yang è trasformata in aminociclopropano-1-carbossilato (ACC) il precursore dell’Et. L’ACC è ossidato dalla ACC ox (ACO) per formare Et, cianuro e CO2.")
38
Regolazione trascrizionale della sintesi dell’Et
ADS è coinvolta in molti metabolismi oltre alla sintesi dell’Et, infatti la sua espressione non viene indotta in modo particolare dagli stress ACO, che è codificata da una famiglia multigenica, è differenzialmente espressa durante lo sviluppo o in risposta all’attacco del patogeno oltre che a stress di natura abiotica. In particolare 2 geni aco (aco1/2) sono upregolati dai fattori di avirulenza AvrPto e AvrPtoB. Quindi la sua espressione è controllata anche a livello trascrizionale ACS è presente in almeno 10 copie geniche in At che hanno un ruolo fondamentale nella sintesi di Et che è infatti regolata attraverso il controllo trascrizionale dei geni acs in risposta a diversi tipi di stimoli. Atacs2 è fortemente upregolato in condizioni di attacco da parte di patogeni necrotrofici come B. cinerea mentre Atacs5 e 11 sono downregolati quanto At è attaccata da biotrofici come P. syringae
 sono upregolati dai fattori di avirulenza AvrPto e AvrPtoB. Quindi la sua espressione è controllata anche a livello trascrizionale. ACS è presente in almeno 10 copie geniche in At che hanno un ruolo fondamentale nella sintesi di Et che è infatti regolata attraverso il controllo trascrizionale dei geni acs in risposta a diversi tipi di stimoli. Atacs2 è fortemente upregolato in condizioni di attacco da parte di patogeni necrotrofici come B. cinerea mentre Atacs5 e 11 sono downregolati quanto At è attaccata da biotrofici come P. syringae.")
39
Percezione e signalling citoplasmico
Et prodotto in seguito a stimolo interno od esterno è percepito dalla cellula e questo segnale è ulteriormente trasmesso attraverso una singola e ben conservata cascata di segnali L'analisi dei mutanti ein2 ha messo in evidenza 5 recettori coinvolti nei primi step di percezione dell'Et, ETR1-2, ERS1-2 e EIN4. Questi hanno una buona omologia con i regolatori a due componenti batterici ETR1 e ERS1 sono presenti nella membrana del reticolo endoplasmatico come omodimeri legati con un ponte disolfuro Ogni omo-dimero forma una tasca idrofobica che rappresenta un sito di legame per l'Et (mediata da un cofattore rameico)
")
40
Percezione e signaling citoplasmico
Altri domini presenti nei recettori ETR sono: un cGMP binding site e una regione HK. Il ruolo di questi domini nella percezione dell'Et ancora non è chiara, anche se la fosforilazione esercitata attraverso un dominio SERK (presente nella subfamiglia 2 dei recettori) è necessaria per trasmettere l'Et signalling a valle Tutti questi recettori per l'Et sono inibiti dal legame con l'ormone e agiscono come regolatori negativi delle risposte all'Et Quindi la percezione dell‘Et avviene a livello dell'ER che rappresenta un punto di giunzione per i diversi processi cellulari che includono diverse risposte allo stress in cui l'Et è un regolatore chiave
 è necessaria per trasmettere l Et signalling a valle. Tutti questi recettori per l Et sono inibiti dal legame con l ormone e agiscono come regolatori negativi delle risposte all Et. Quindi la percezione dell‘Et avviene a livello dell ER che rappresenta un punto di giunzione per i diversi processi cellulari che includono diverse risposte allo stress in cui l Et è un regolatore chiave.")
41
Percezione e signaling citoplasmico
Studi sul mutante ctr1 (constitutive triple response) di At hanno portato all'individuazione di CTR1 come regolatore negativo delle risposte all'Et. Oltre ad avere un dominio a funzione sconosciuta nell'N terminale possiede un dominio SERK al C terminale (che lo rende simile alle Rak-like kinases) Anche CTR1 si trova nell'ER anche se non ha nessun segnale di riconoscimento per la membrana ma la sua localizzazione è suggerita dall'interazione con i recettori per l'Et. Sembra che sia l'interazione con i recettori che l'attività kinasica siano necessari per la regolazione negativa delle risposte all'Et
 di At hanno portato all individuazione di CTR1 come regolatore negativo delle risposte all Et. Oltre ad avere un dominio a funzione sconosciuta nell N terminale possiede un dominio SERK al C terminale (che lo rende simile alle Rak-like kinases) Anche CTR1 si trova nell ER anche se non ha nessun segnale di riconoscimento per la membrana ma la sua localizzazione è suggerita dall interazione con i recettori per l Et. Sembra che sia l interazione con i recettori che l attività kinasica siano necessari per la regolazione negativa delle risposte all Et.")
42
Percezione e signaling citoplasmico
In assenza di Et il legame con i recettori mantiene CTR1 attivo agendo cosi come regolatore negativo abile nel reprimere le risposte a valle indotte dall'Et. Il legame dell'Et con i recettori causa un cambiamento conformazionale di CTR1 che risulta nel rilascio del suo effetto inibitorio Un altro mutante risultante dallo screening per l'insensibilità all'Et è risultato essere ein2. EIN2 possiede due domini, uno idrofilico che è coinvolto nell'interazione P-P e uno idrofobico che contiene 12 eliche transmebrana che la rendono simile alle Nramp proteins coinvolte nel trasporto transmembrana dei metalli

43
Trasmissione nucleare dell'Et signal
La completa perdita di sensibilità all'Et nei mutanti ein2 indica un ruolo centrale nella trasduzione del segnale da parte di EIN2. L'overespressione della parte C terminale sembra essere sufficiente ad attivare in modo costitutivo le risposte all'Et Quindi il segnale dell'Et arriva nel nucleo attraverso la derepressione di EIN2 da parte di CTR1 e porta all'attivazione dei fattori di trascrizione EIN3 e EIN3-like EIN3 ed EIL1 sono correlati tra loro e sono i fattori più importanti nella sensibilità all'Et La modulazione dell'attività di questi fattori dall'Et non avviene a livello trascrizionale ma attraverso il controllo proteolitico dei livelli di EIN3 da parte del complesso SCF ubiquitina ligasi

44
Trasmissione nucleare dell'Et signal
In assenza di Et EIN3 sono continuamente degradate attraverso AtEBF1 (F-box protein che destabilizzano le proteine target) e il proteasoma e in tale modo si previene l'attivazione dei loro target trascrizionali. Mentre, in presenza di Et la degradazione di EIN3 è soppressa consentendo così a questo fattore di incrementarsi in quantità e di promuovere gli eventi a valle Dato che diverse PK sono poste a monte di EIN3 è possibile che la sua fosforilazione inibisca l'associazione con AtEBF1 prevenendone quindi la sua degradazione EIN3 riconosce il suo target nel DNA chiamato EIN3 binding site (EBS) o primary Et response element (PERE) che si trova nei promotori dei geni che codificano per proteine che si legano negli elementi di risposta all'Et
 e il proteasoma e in tale modo si previene l attivazione dei loro target trascrizionali. Mentre, in presenza di Et la degradazione di EIN3 è soppressa consentendo così a questo fattore di incrementarsi in quantità e di promuovere gli eventi a valle. Dato che diverse PK sono poste a monte di EIN3 è possibile che la sua fosforilazione inibisca l associazione con AtEBF1 prevenendone quindi la sua degradazione. EIN3 riconosce il suo target nel DNA chiamato EIN3 binding site (EBS) o primary Et response element (PERE) che si trova nei promotori dei geni che codificano per proteine che si legano negli elementi di risposta all Et.")
45
Trasmissione nucleare dell'Et signal
Il fattore di trascrizione ERF1 è un immediato target per EIN3. ERF1 appartiene alla famiglia dei fattori di risposta all'Et anche chiamati proteine che si legano agli elementi di risposta all'Et (EREBs). Questi fattori di trascrizione si legano nel cosiddetto GCC box che è un elemento presente nel promotore di molti geni responsive all'Et come ad es. quelli che codificano per le PR proteins Il GCC box è un elemento che agisce in cis e che consiste di una sequenza conservata di 11 bp (TAAGAGCCGCC) necessaria e sufficiente per la regolazione da parte dell'Et dei geni responsive Gli ERF mostrano un'alta omologia con i fattori di trascrizione che si legano ai siti DREB (dehydration responsive element) nei promotori dei geni che sono responsive allo stress abiotico
. Questi fattori di trascrizione si legano nel cosiddetto GCC box che è un elemento presente nel promotore di molti geni responsive all Et come ad es. quelli che codificano per le PR proteins. Il GCC box è un elemento che agisce in cis e che consiste di una sequenza conservata di 11 bp (TAAGAGCCGCC) necessaria e sufficiente per la regolazione da parte dell Et dei geni responsive. Gli ERF mostrano un alta omologia con i fattori di trascrizione che si legano ai siti DREB (dehydration responsive element) nei promotori dei geni che sono responsive allo stress abiotico.")
46
Trasmissione nucleare dell'Et signal
La superfamiglia AP/ERF di fattori di trascrizione comprende circa 145 membri in At In aggiunta alla regolazione trascrizionale attraverso i fattori EIN3, i fattori Erf sono regolati a livello post mediante la fosforilazione, infatti AtERF5 contiene un sito riconosciuto da una MAPK e la sua fosforilazione aumenta la sua capacità di legame ai GCC box Molti ERF sono degli attivatori trascrizionali come AtERF1, 2 e 5 mentre una seconda classe di proteine ERF come AtERF3, 4, 7 e 10 reprimono l'espressione dei geni che presentano un GCC box In tutti gli ERF è presente un motivo AA di repressione genica chiamato EAR (ERF associated amphiphilic repression). L'overespressione di ERF4 conferisce un fenotipo Et insensibile e reprime l'espressione di tutti i geni contenenti nel promotore un GCC box
. L overespressione di ERF4 conferisce un fenotipo Et insensibile e reprime l espressione di tutti i geni contenenti nel promotore un GCC box.")
47
Etilene nella malattia
La biosintesi di Et è attivata in molte piante in seguito ad attacco patogenico e la sua produzione induce dei geni effettori relativi alla difesa attraverso una cascata di eventi di cui il penultimo step è rappresentato dall'attivazione dei fattori di trascrizione ERF-like Il ruolo specifico dell'Et nella difesa è centrato su 3 fronti poiché induce Proteine che partecipano nel build-up delle barriere fisiche Enzimi del metabolismo secondario coinvolti nella biosintesi delle fitoalessine Proteine relative alla patogenesi (PR)
")
48
Cross talk ormonale

Presentazioni simili

















 Le cellule distanti tra loro comunicano attraverso molecole (MEDIATORI o NEUROTRASMETTITORI)>")



