Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
CORSO DI LAUREA SPECIALISTICA IN BIOTECNOLOGIE DEL FARMACO
Adriana Maggi BIOTECNOLOGIE FARMACOLOGICHE LEZIONE 14

2
FARMACI BIOTECNOLOGICI
PROTEINE TERAPEUTICHE DI PRIMA E SECONDA GENERAZIONE ACIDI NUCLEICI per la regolazione della espressione di specifici geni

3
VANTAGGI NELL’UTILIZZO DI FARMACI
ANTI-ACIDI NUCLEICI BERSAGLIO MOLECOLARE PIU’ DEFINITO Numero di proteine intracellulari: Numero di RNA: Numero di geni: 1-2 AZIONE MOLTO SPECIFICA Bersagli possono essere sequenze anche uniche nel genoma Utilizzando interazioni con altri nucleotidi si possono sfruttare i vantaggi della complementarità tra due catene secondo il modello di Watson e Crick

4
…IL MEDICO ADOPRERA’ BENE
LE MEDICINE QUANDO LUI CONOSCERA’ CHE COSA E’ OMO, CHE COSA E’ VITA E COMPLESSIONE, CHE COSA E’ SANITA’…

5
ESPRESSIONE GENICA IN EUCARIOTE: SEQUENZA DI EVENTI
3’ TRASCRIZIONE MODIFICAZ. TP TRADUZIONE RNA, trascritto primario Capping e poliadenilazione Splicing o maturazione Fuoriuscita dal nucleo riconoscimento da parte dei ribosomi traduzione Modificazioni post-traduzionali

6
La rilevanza dei processi di regolazione della espressione genica
nel disegno di nuovi farmaci L’omeostasi cellulare è mantenuta da un efficiente controllo della trascrizione genica che viene assicurato da un complesso circuito molecolare che utilizza fattori di trascrizione individali, l’apparato basale di trascrizione e i complessi multiproteici coregolatori della trascrizione.

7
La cromatina ha un ruolo fondamentale nella reglazione della espressione genica e la capacità degli istoni di legare il DNA è regolata da enzimi intranucleari che possono apportare modificazioni post-traduzionali soprattutto ai residui di lisina e arginna posti nel dominio N-terminale delle proteine istoniche Modificazioni degli istoni: acetilazione, fosforilazione, metilazione, ubiquitinazione

8
MODIFICAZIONI POST-TRADUZIONALI DEGLI ISTONI
Zhang Y , Reinberg D Genes Dev. 2001;15:

9
Metilazione degli istoni
La metilazione degli istoni aumenta lo stato di idrofobicità e la basicità delle porzioni N trminali degli istoni: questo ne aumenta l’affinità per proteine specifiche e fattori di trascrizione. Le metilazsi necessitano di cofattori: es. la s-adenosilmetionina Generalmente la metilazione egli istoni si associa a repressione della espressione genica, tuttavia ci sono esempi in cui la metilazione si associa a attivazione della espressione genica (metilazione lella lisina 4 dell’istone 3, il coattivatore CARM1 è una metiltransferasi)
")
10
PROCESSI DI ACETILAZIONE E
DEACETILAZIONE SONO IMPORTANTI NELLA REGOLAZIONE DELLA ATTIVITA’ DELLA CROMATINA ESPRESSIONE GENICA E’ GENERALMENTE ASSOCIATA CON ACETILAZIONE

11
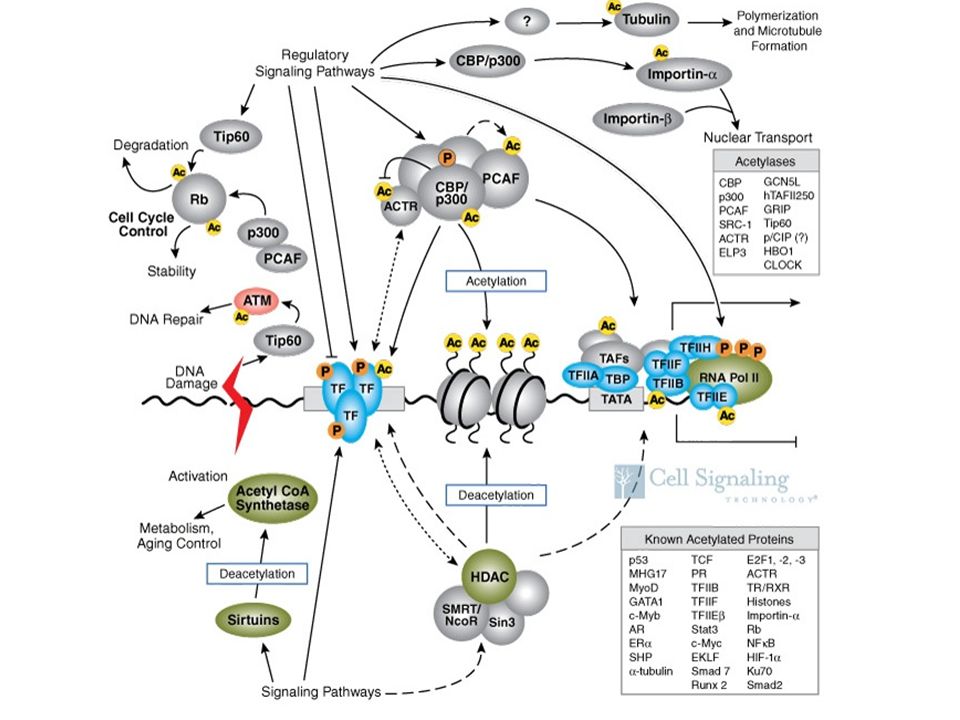
L’acetilazione dei residui di lisina al terminale NH degli istoni rimuove le cariche positive riducendo quindi l’affinità degli istoni per il DNA: per questo l’acetilazione degli istoni facilita i processi trascrizionali , al ontrario, la deacetilazione reprime la trascrizione . Acetilazione e deacetilazione del residuo lisinico ACETIL TRANSFERASI ISTONICHE :histone acetyltransferases (HATs) DEACETILASI ISTONICHE: histone deacetylases (denoted by HDs or HDACs); numerose deacetilasi sono associate a repressori della trascrizione;
 DEACETILASI ISTONICHE: histone deacetylases (denoted by HDs or HDACs); numerose deacetilasi sono associate a repressori della trascrizione;")
12
Fosforilazione degli istoni
la funzione della fosforilazione degli istoni rimane da essere pienamente compresa: è stato riportato che la fosforilazione degli istoni puo’ facilitare l’attacco di HATs Enzimi quali le ERK possono fosforilare gli istoni legando signaling di recettori di membrana allo stato di fosforilazione degli istoni

13
Fattori di rimodellamento della cromatina
Due famiglie di fattori sono coinvolte nel rimodellamento della cromatna: SWI/SFI e ISWI: in genere queste proteine destabilizzano il legame tra istoni e DNA aiutando lo “scivolamento” dei nucleosomi e la trascrizione del DNA : queste proteine fungono quindi da enhancers della trascrizione. Esistono inoltre proteine non istoniche che facilitano il legame delle HAT Altri fattori di rimodellamento della cromatina agiscono utilizzando ATP e possono riposizionare I nucleosomi sulla cromatina (ACF e NAP-1)
")
14
Meccanismi di metilazione del DNA e eredità epigenetica

15
Meccanismi di metilazione del DNA che agiscono su sequenze palindromi:
e eredità epigenetica Circa il 2-7% delle citosine nel DNA animale subisce metilazione ad opera di specifici enzimi che agiscono su sequenze palindromi: 5’ mCpG 3’ 3’ GpCm 5’

16
CpG islands are genomic regions that contain a high frequency of CG nucleotides. In mammalian genomes, CpG islands are typically 300-3,000 base pairs in length. They are in and near approximately 40% of promoters of mammalian genes (about 70% in human promoters). The "p" in CpG notation refers to the phosphodiester bond between the cytosine and the guanine. The usual formal definition of a CpG island is a region with at least 200 bp and with a GC percentage that is greater than 50% and with an observed/expected CpG ratio that is greater than 60%. Another recent study revised the rules of CpG island prediction in order to exclude other GC-rich genomic sequences such as Alu repeats. Based on an extensive search on the complete sequences of human chromosomes 21 and 22, DNA regions >500 bp with a GC content >55% and observed CpG/expected CpG of 0.65 were more likely to be the true CpG islands associated with the 5' regions of genes.

17
ESPRESSIONE GENICA E’ GENERALMENTE
ASSOCIATA CON DEMETILAZIONE I GENI “HOUSEKEEPING” SONO ESPRESSI IN MODO COSTITUTIVO E CONTENGONO NEL LORO PROMOTORE MOLTE “ISOLE” CpG NON METILATE

18
A Unifying Model for the Selective Regulation of Inducible Transcription by CpG Islands and Nucleosome Remodeling Vladimir R. Ramirez-Carrozzi,1,2,5 Daniel Braas,1,2,5 Dev M. Bhatt,1,2,5 Christine S. Cheng,4 Christine Hong,1,2 Kevin R. Doty,1,2 Joshua C. Black,2,3 Alexander Hoffmann,4 Michael Carey,2,3 and Stephen T. Smale1,2,* Cell 138: 114, 2009 CpG island promoter: 70% dei promotori hanno numerose sequenze ricche in CpG : basso livello di metilazione delle CpG island che li rende indipendenti da SWI/SNF remodeling complexes Non CpG island promoter: 30% dei promotori che sono altamente regolati trascriionalmente

19
Sindrome di Rett MeCP2 gene

20
methyl DNA binding protein 2 (MeCP2)
The mechanism(s) by which the native MeCP2 protein operates in the cell are not well understood. Historically, MeCP2 has been characterized as a proximal gene silencer with 2 functional domains: a methyl DNA binding domain and a transcription repression domain. However, several lines of new data indicate that MeCP2 structure and function relationships are more complex: an analysis of cell-based experiments suggesting MeCP2 is a regulator, rather than a strict silencer, of transcription. The new data establish MeCP2 as a multifunctional nuclear protein, with potentially important roles in chromatin architecture, regulation of RNA splicing, and active transcription..
 by which the native MeCP2 protein operates in the cell are not well understood. Historically, MeCP2 has been characterized as a proximal gene silencer with 2 functional domains: a methyl DNA binding domain and. a transcription repression domain. However, several lines of new data indicate that MeCP2 structure and function relationships are more complex: an analysis of cell-based experiments suggesting MeCP2 is a regulator, rather than a strict silencer, of transcription. The new data establish MeCP2 as a multifunctional nuclear protein, with potentially important roles in chromatin architecture, regulation of RNA splicing, and active transcription..")
21
methyl DNA binding protein 2 (MeCP2)
MeCP2 binds preferentially to DNA methylated at CpG sites through an 85–amino acid methyl-CpG–binding domain (MBD) (amino acids 78–162) Once bound to DNA, MeCP2 is thought to silence transcription of downstream genes by recruiting corepressor complexes through a 104–amino acid transcriptional repression domain (TRD) (amino acids 207–310) One complex shown to associate with MeCP2 is the Sin3A corepressor complex. This complex contains histone deacetylase (HDAC) 1 and HDAC2 and was originally shown to mediate repression by the DNA-binding heterodimer, Mad-Max . MeCP2 also interacts with two other corepressors, the proto-oncoprotein of the Sloan-Kettering virus named after the Sloan-Kettering Institute (c-Ski) and the nuclear receptor corepressor (N-CoR). c-Ski and N-CoR are components of histone deacetylase complexes that can but do not always function together It is interesting that repression by MeCP2 is not completely alleviated by the histone-deacetylase inhibitor, trichostatin A (TSA), suggesting that MeCP2 may also repress transcription in an HDAC-independent manner
 (amino acids 78–162) Once bound to DNA, MeCP2 is thought to silence transcription of downstream genes by recruiting corepressor complexes through a 104–amino acid transcriptional repression domain (TRD) (amino acids 207–310) One complex shown to associate with MeCP2 is the Sin3A corepressor complex. This complex contains histone deacetylase (HDAC) 1 and HDAC2 and was originally shown to mediate repression by the DNA-binding heterodimer, Mad-Max . MeCP2 also interacts with two other corepressors, the proto-oncoprotein of the Sloan-Kettering virus named after the Sloan-Kettering Institute (c-Ski) and the nuclear receptor corepressor (N-CoR). c-Ski and N-CoR are components of histone deacetylase complexes that can but do not always function together. It is interesting that repression by MeCP2 is not completely alleviated by the histone-deacetylase inhibitor, trichostatin A (TSA), suggesting that MeCP2 may also repress transcription in an HDAC-independent manner.")
22
Modeling Rett Syndrome with Stem Cells

23
METILAZIONE DEL DNA E CANCRO
The alterations of DNA methylation level and patterns are a common feature of human cancer cells. A global DNA hypomethylation has been observed in many cancers, without obvious sequence specificity Recently, an extensive study of about 1200 CpG islands has indicated that hypermethylated CpG islands are not randomly distributed and the patterns of the hypermethylation might be specific of subclasses of cancers. The methylation status of tumor suppressor genes has been extensively investigated and such alterations have been reported in many human tumors (Robertson and Jones, 2000).
.")
24
METILAZIONE DEL DNA E CANCRO
Several reports link genome hypomethylation to genome instability. In particular, it was shown recently that strongly reduced DNMT activity in a transgenic mouse model caused demethylation of centromeric satellite and other repeat sequences, which resulted in a variety of chromosome defects and concomitant tumorigenesis

25
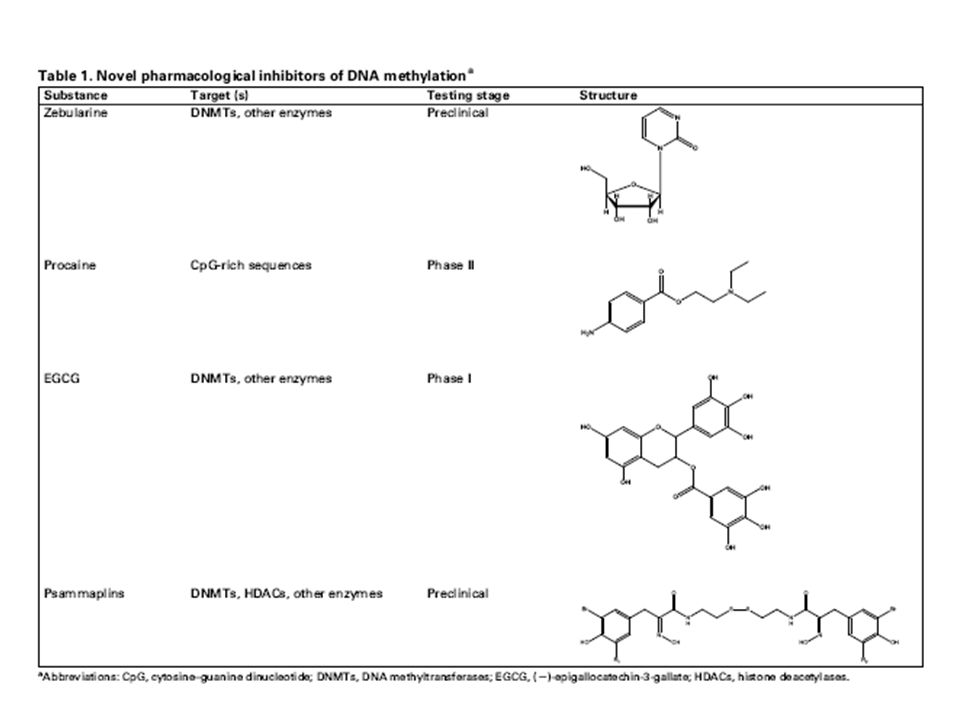
Peter Jones nel 1970 descrisse il differenziamento di cellule embrioniche in cellule muscolari dopo esposizione a una molecola capace di aumentare lo stato di metilazione del DNA (5-Azacytidine) . Nel 2004 la 5-azacytidine ( Vidaza) è stato approvato per il trattamento della sindrome mielodisplastica Nel 2006 è stato approvata un’altra molecola la decitabina (Dacogen) : queste molecole sono substrati dell DNA methyltransferasi. Altri farmaci epigenetici in studio sono gli inibitori delle acetiltransferasi (HDAC inhibitors) il primo di questi, SAHA (zolinza) è stato approvato nel 2006 per il linfoma a cellule T cutaneo.
 è stato approvato per il trattamento della sindrome mielodisplastica. Nel 2006 è stato approvata un’altra molecola la decitabina (Dacogen) : queste molecole sono substrati dell DNA methyltransferasi. Altri farmaci epigenetici in studio sono gli inibitori delle acetiltransferasi (HDAC inhibitors) il primo di questi, SAHA (zolinza) è stato approvato nel 2006 per il linfoma a cellule T cutaneo.")
26
VORINOSTAT (Zolinza) è il primo inibitore delle acetilasi istoniche approvato per il trattamento di neoplasie Nel 2006 il VORINOSTAT è stato approvato per il trattamento del linfoma a cellule T cutaneo Esistono studi preclinici che indicano una attività antinfiammatoria del vorinostat Nel 2007 ricerche presso la Mayo Clinics hanno dimostrato che il Vorinostat è efficace nel glioblastoma ricorrente N H O OH N-hydroxy-N'-phenyl-octanediamide

27
HDAC inhibitors can activate both the death-receptor and intrinsic apoptotic pathways

29
A LIVELLO TRASCRIZIONALE
REGOLAZIONE DELLA ESPRESSIONE GENICA IN EUCARIOTE A LIVELLO TRASCRIZIONALE

30
TFII H helicase and kinase
TBP + 12 TAFs (TBP ass. factors) TFIID recognizes TATA (Inr) commitment TFIIA; TFIIB; TFIIF Pol II entry TFIIE; TFIIH Promoter clearence TFIIF pol II binding TFII H helicase and kinase Bob Roeder Lasker Award 2003
 TFIID recognizes TATA (Inr) commitment. TFIIA; TFIIB; TFIIF. Pol II entry. TFIIE; TFIIH. Promoter clearence. TFIIF pol II binding. TFII H helicase and kinase. Bob Roeder. Lasker Award")
31
Promotori prossimali e promotori distali

33
ESPRESSIONE GENICA IN EUCARIOTE: SEQUENZA DI EVENTI
3’ TRASCRIZIONE MODIFICAZ. TP TRADUZIONE RNA, trascritto primario Capping e poliadenilazione Splicing o maturazione Fuoriuscita dal nucleo riconoscimento da parte dei ribosomi traduzione Modificazioni post-traduzionali

34
Biogenesi di RNA messaggero e regolazione genica post-trascrizionale
Cell 136:777, 2009

35
INTRONI DI RNA NUCLEARE BERSAGLIO DELL’APPARATO DI SPLICING

36
Ruolo snRNP e proteine associate nello splicing 1
5’ 3’ Commitment complex (E complex) U1 lega il sito di taglio in 5’ e U2AF lega il sito polipirimidinico 5’ 3’ 5’ 3’ Complex A U2 lega il sito “branch” Complex B si associano U5, U6, U4
 U1 lega il sito di taglio in 5’ e U2AF lega. il sito polipirimidinico. 5’ 3’ 5’ 3’ Complex A U2 lega il sito branch Complex B si associano U5, U6, U4.")
37
Ruolo snRNP e proteine associate nello splicing 2.
Riposizionamento, U1 si dissocia, U5 si associa all’introne; U 6 lega il sito di taglio in 5’ Complex C U4 si dissocia, U6 e U2 catalizzano la transesterificazione U5 l’esone al sito di taglio in 3’ Avviene il taglio in 5’ e si forma il lariate Avviene il taglio in 3’ e la ligazione Tra gli esoni

38
LE SEQUENZE DI SPLICING SONO CONSERVATE NEI VERTEBRATI
5’ splice site: 5’-GU-3’ Tratto polipirimidinico 3’ splice site 5’-AG-3’

39
Esistono introni AU-AC sono simili a introni GU-AG ma richiedono
Un apparato di splicing differente

40
L’eliminazione di sequenze di RNA che non abbiano subito il processamento è assicurata dalla presenza di “nonsense-mediated mRNA decay” un meccanismo di turnover di RNA che degrada sequenze in cui sia presente un segnale di stop Questo suggerisce la co-evoluzione dei meccanismi di splicing e di traduzione del segnale

41
Mutazioni che alterano il codice di slicing causano patologie specifiche

42
Cell 136: 777, 2009

43
Micro RNA e regolazione della espressione genica

44
ACIDI NUCLEICI IN TERAPIA
ANTISENSO: desossinucleotidi complementari con l’m-RNA ANTIGENE: desossinucleotidi complementari al DNA RIBOZIMA: ribonucleotidi catalalitici complementari a RNA APTAMERI: desossi e ribonucleotidi complementari a una sequenza aminoacidica (competono con l’RNA nell’interazione tra RNA e proteina) DECOY: DNA complementare a sequenze aminoacidiche (competono con sequenze di DNA nell’interazione DNA-proteina) siRNA: RNA complementari a sequenze specifiche
 DECOY: DNA complementare a sequenze aminoacidiche (competono con sequenze di DNA nell’interazione DNA-proteina) siRNA: RNA complementari a sequenze specifiche.")
45
Nuovi target farmacologici: GLI ACIDI NUCLEICI
PROTEINA FARMACI TRADIZIONALI DNA duplicato DNA RNA Duplicazione Trascrizione Maturazione Metabolismo Traduzione

Presentazioni simili


















 Piccole molecole di RNA (20-22 nt)>")
