Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

2
Lo stomaco deve operare in maniera tale da consentire all’intestino di svolgere in maniera ottimale le funzioni di digestione ed assorbimento. Questo avviene attraverso diversi processi che sono:

3
Distensione della parete gastrica con riduzione della pressione endogastrica quando il bolo arriva allo SEI Accomodamento della tensione della parete addominale al volume gastrico. Lo stomaco adatta la sua distensione al volume del contenuto. Omogeneizzazione e rimescolamento dei componenti Svuotamento controllato e selettivo e svuotamento delle particelle non digeribili Regolazione del reflusso duodeno-gastrico

4
Le principali funzioni possono essere quindi così riassunte 1.Storage di grandi quantità di cibo che saranno poi processate nel duodeno 2.Mescolamento del cibo con le secrezioni gastriche fino a formare una poltiglia acida detta chimo 3.Lento svuotamento dallo stomaco al duodeno con una rate adatta ad un’appropriata digestione ed assorbimento

5
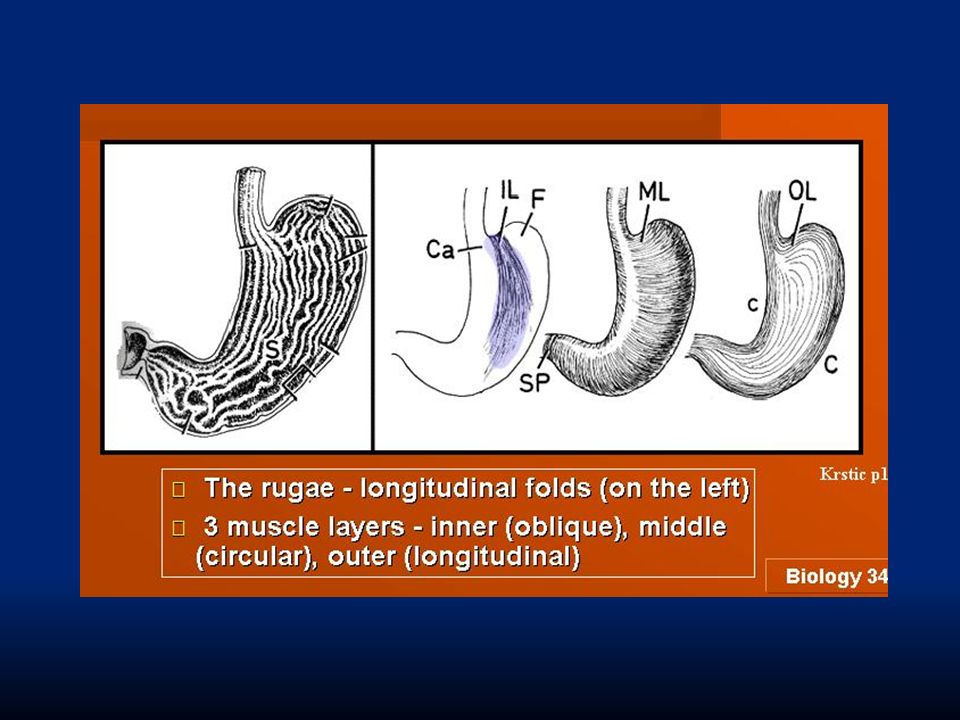
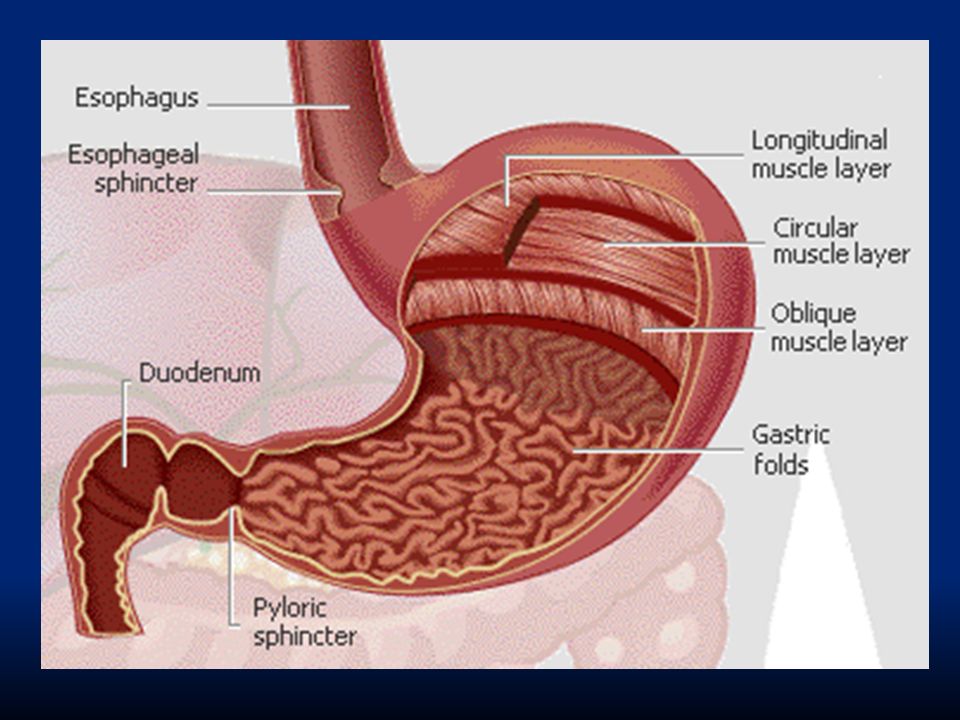
Lo stomaco è localizzato sul lato sinistro del corpo, al di sotto del diaframma. Connette esofago ed intestino ed è un organo sacciforme, caratterizzato da un triplo strato muscolare (longitudinale, circolare ed obliquo) che garantisce un movimento notevole per mescolare il cibo. longitudinale(esterno)circolare(medio)obliquo(interno)
 che garantisce un movimento notevole per mescolare il cibo. longitudinale(esterno)circolare(medio)obliquo(interno).")
8
A livello di stomaco troviamo una porzione che corrisponde alla grande curva detta fondo, il corpo, e un’ altra parte detta antro.

9
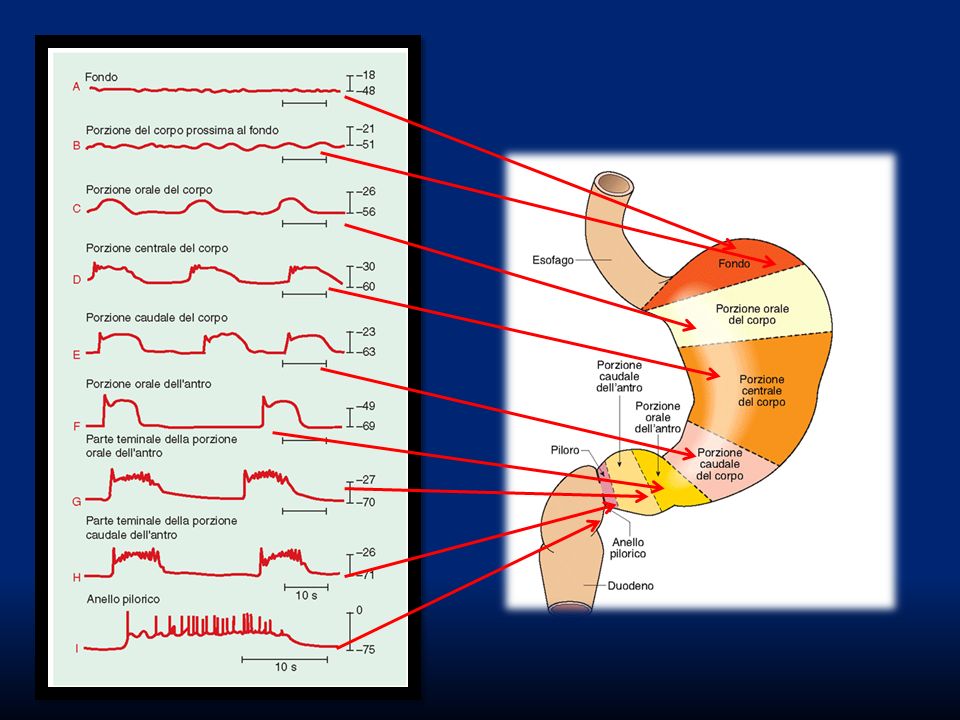
antro Zona prossimale= serbatoio Zona antrale o distale= contrazioni ritmiche e peristaltiche Zona pilorica= regola lo svuotamento e il reflusso

10
Storage: quando il cibo entra nello stomaco forma anelli concentrici con il cibo più recente in prossimità dello sfintere esofageo e quello più vecchio addossato alle pareti dello stomaco. Un riflesso vago - vagale evocato dalla distensione dell’organo provoca un rilasciamento della parte dell’organo in modo che possa accomodare quantità crescenti di cibo.

11
Si distinguono tre diversi periodi con diversa attività elettrica e meccanica: Periodo interdigestivo: attività ciclica detta complesso mioelettrico migrante (CMM) che alterna momenti di intensa attività a momenti quiescenti. In particolare la fase 3 presenta spike intensi e regolari con contrazioni intense (crampi da fame). Lo stimolo prende origine dal pacemaker gastrico.
. Lo stimolo prende origine dal pacemaker gastrico..")
12
Arrivo del bolo nello stomaco: quando il bolo arriva nello stomaco il CMM ha termine. Si assiste al rilasciamento recettivo di cardias e fondo dello stomaco per accogliere il cibo. Questo è mediato da recettori esofagei sensibili alla distensione e da fibre vagali. Periodo digestivo: la presenza di cibo nello stomaco determina per riflessi locali e vagali un progressivo aumento della motilità gastrica che non è uniforme ma va aumentando verso l’antro e il piloro

14
LES Fondo Scarsa attività elettrica basale Scarsa attività elettrica basale Contrazioni toniche lente Contrazioni toniche lente Elevata distensibilità Elevata distensibilità Reservoir gastrico Reservoir gastrico Corpo e antro Attività elettrica pronunciata Attività elettrica pronunciata Contrazioni fasiche peristaltiche Contrazioni fasiche peristaltiche Scarsa distensibilità Scarsa distensibilità Sminuzzamento di particellle solide Sminuzzamento di particellle solide

15
Mescolamento: appena il cibo è nello stomaco le oscillazioni lente spontanee determinano un aumento dei movimenti di mescolamento da metà circa dello stomaco in direzione dell’antro. Verso l’antro queste onde divengono più potenti formando anelli peristaltici che spingono il chimo verso il piloro. Quando questo si apre fa passare pochi ml di chimo per volta e poi si richiude generando un movimento di retropulsione.

16
Movimento di retropulsione nell’antro

17
Il volume dello stomaco in seguito ad un pasto può arrivare a circa 1.5 litri

18
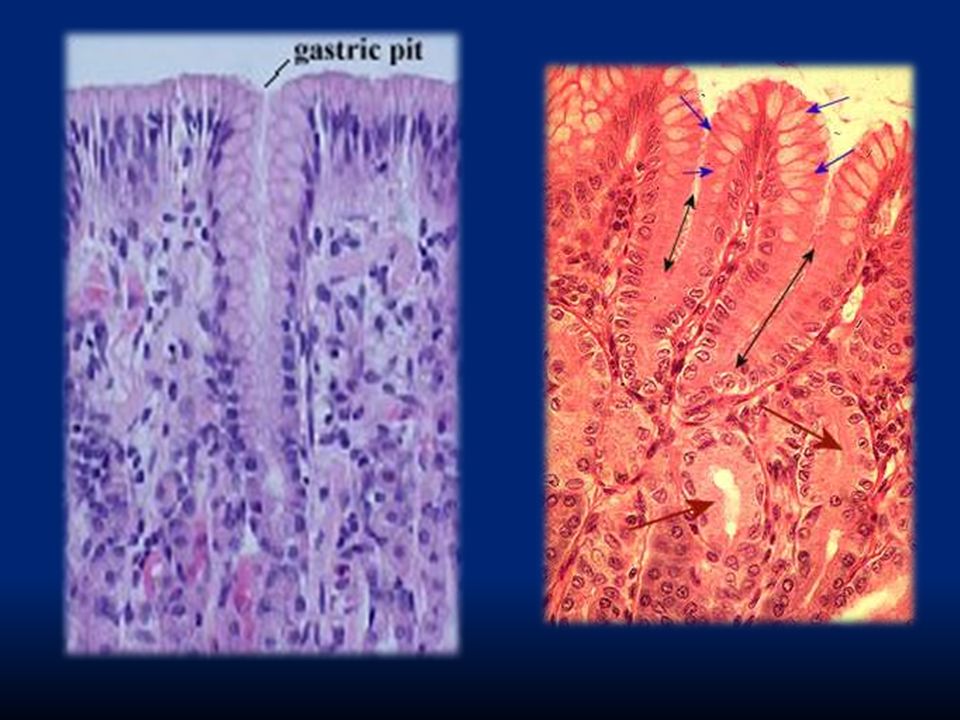
Epitelio colonnare di secrezione

19
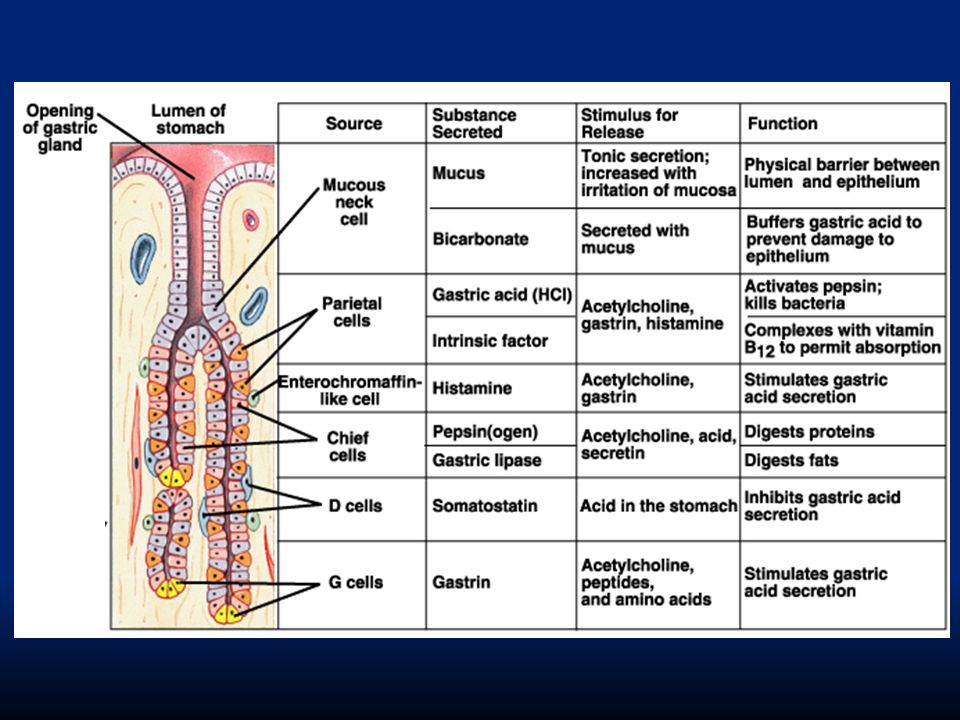
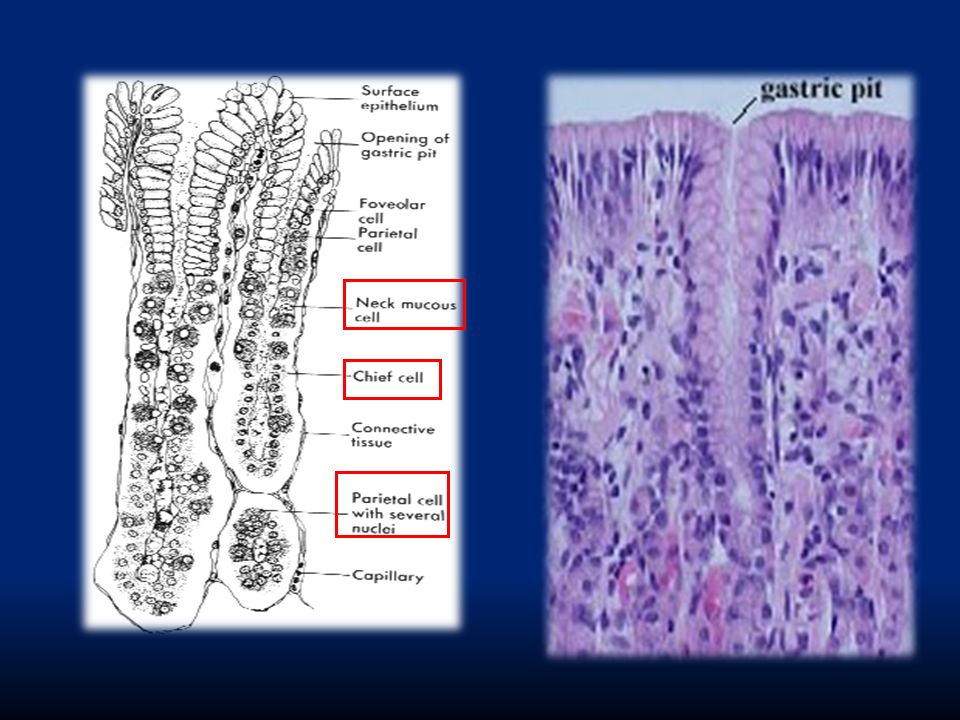
Lo strato epiteliale è formato da un epitelio colonnare monostratificato interrotto da “fori” che corrispondono all’apertura di ghiandole gastriche (gastric pits). Troveremo quattro diversi tipi di secrezione: Cellule mucose: secernono muco alcalino per formare la barriera gastrointestinale Cellule parietali: secernono HCl Cellule principali: secernono pepsina Cellule G: secernono gastrina

21
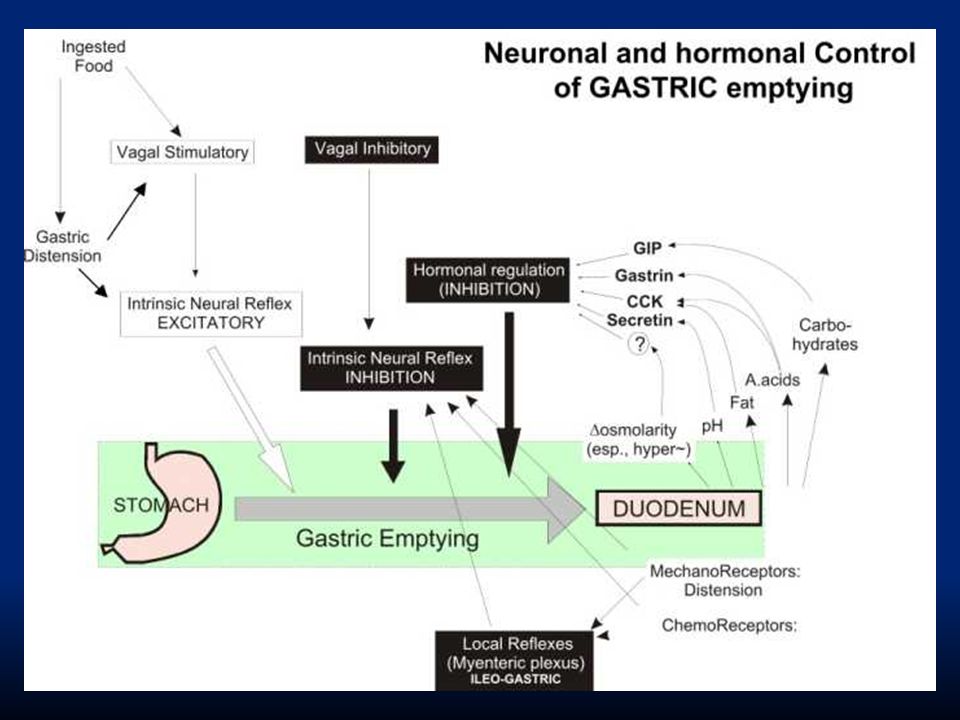
Lo svuotamento gastrico è promosso dalle intense contrazioni dell’antro. Queste sono contrazioni a forma di anello che generano un’elevata pressione e determinano svuotamento del chimo acido nel duodeno fornendo un’azione di pompa indicata con il nome di “pompa pilorica”. Esistono anche vari impedimenti allo svuotamento dello stomaco, che vedremo successivamente.

22
Il piloro è lo sfintere che controlla l’uscita dello stomaco ed è dotato di una muscolatura molto forte che rimane tonicamente contratta: si parla di sfintere pilorico. Il piloro si apre per permettere il passaggio di acqua e liquidi, ma in genere non di particelle che debbano ancora essere ridotte a chimo. Quali sono i fattori che favoriscono lo svuotamento gastrico e quelli che inibiscono?

23
Contrazioni peristaltiche dell’antro dell’antro Aumento del volume di cibo nello stomaco (riflessi enterici evocati dallo stiramento della parete determinano aumento della peristalsi) Gastrina rilasciata dalle cellule dell’antro (stimola la pompa pilorica) Gastrina rilasciata dalle cellule dell’antro (stimola la pompa pilorica) Svuotamento gastrico Fattori gastrici stimolatori
 Gastrina rilasciata dalle cellule dell’antro (stimola la pompa pilorica) Gastrina rilasciata dalle cellule dell’antro (stimola la pompa pilorica) Svuotamento gastrico Fattori gastrici stimolatori")
24
Svuotamento gastrico Effetto inibitorio di riflessi nervosi enterogastrici * CCK in risposta a sostanze grasse nel duodeno nel duodeno Secretina e GIP rispettivamente in risposta a grado di acidità e carboidrati / grassi Fattori inibitori duodenali

25
I riflessi inibitori hanno tre origini: * I riflessi inibitori hanno tre origini: direttamente dal duodeno allo stomaco attraverso il SNE attraverso afferenze simpatiche ai gangli paravertebrali e da qui indietro allo stomaco attraverso il nervo vago al tronco encefalico dove inibiscono i normali segnali eccitatori allo stomaco

26
I fattori che stimolano questi tipi di riflessi sono: distensione della parete duodenale irritazione della mucosa duodenale grado di acidità del chimo grado di osmolarità del chimo presenza di prodotti di degradazione delle proteine e dei grassi Lo svuotamento è inoltre inibito da certi ormoni come la CCK, rilasciata dalla mucosa del digiuno in presenza di sostanze grasse. Lo stesso vale per la secretina.

28
Riflesso del vomito espulsione del contenuto gastrico (e duodenale) attraverso la bocca per antiperistalsi che si origina dal duodeno, stomaco e si propagano all’esofago preceduto da: nausea, tachicardia, vertigini, sudorazione, pallore e dilatazione pupillare preceduto da “conati” atto riflesso controllato da centri bulbari regioni periferiche influenzano il c. del vomito varie sostanze chimiche inducono vomito (emetici); possono agire centralmente o a livello gastrico
; possono agire centralmente o a livello gastrico.")
29
Il vomito è il risultato di un complesso meccanismo, regolato da una ''centralina di comando'' collocata in una particolare zona del cervello. A questo ''centro del vomito'' arrivano segnali da varie parti dell'organismo: dalla corteccia cerebrale, che può stimolare il centro mandando messaggi di sapori, odori, ricordi, emozioni, immagini dalla corteccia cerebrale, che può stimolare il centro mandando messaggi di sapori, odori, ricordi, emozioni, immagini dall'orecchio, che trasmette soprattutto informazioni relative alla posizione del corpo rispetto allo spazio (chinetosi) dall'orecchio, che trasmette soprattutto informazioni relative alla posizione del corpo rispetto allo spazio (chinetosi) da diverse parti dell'apparato digerente (gola, stomaco, intestino) e da altri organi, come il cuore. da diverse parti dell'apparato digerente (gola, stomaco, intestino) e da altri organi, come il cuore.
 dall orecchio, che trasmette soprattutto informazioni relative alla posizione del corpo rispetto allo spazio (chinetosi) da diverse parti dell apparato digerente (gola, stomaco, intestino) e da altri organi, come il cuore. da diverse parti dell apparato digerente (gola, stomaco, intestino) e da altri organi, come il cuore..")
30
Sensibile a stimoli chimici e acidosi bulbare

31
I movimenti dell’intestino tenue comprendono movimenti peristaltici e movimenti di mescolamento che hanno le seguenti funzioni: ►Miscelare i prodotti alimentari con i succhi digestivi ►Facilitare il contatto fra mucosa e chimo ►Far avanzare il contenuto intestinale

32
Distinguiamo una motilità della fase interdigestiva (A) e una motilità della fase digestiva (B). A. L’intestino è attraversato dal complesso mioelettrico migrante (CMM) che origina dal pacemaker gastrico. Onde peristaltiche che interessano corti segmenti intestinali. Il contenuto intestinale è spinto verso la valvola ileo-cecale.
 che origina dal pacemaker gastrico. Onde peristaltiche che interessano corti segmenti intestinali. Il contenuto intestinale è spinto verso la valvola ileo-cecale..")
33
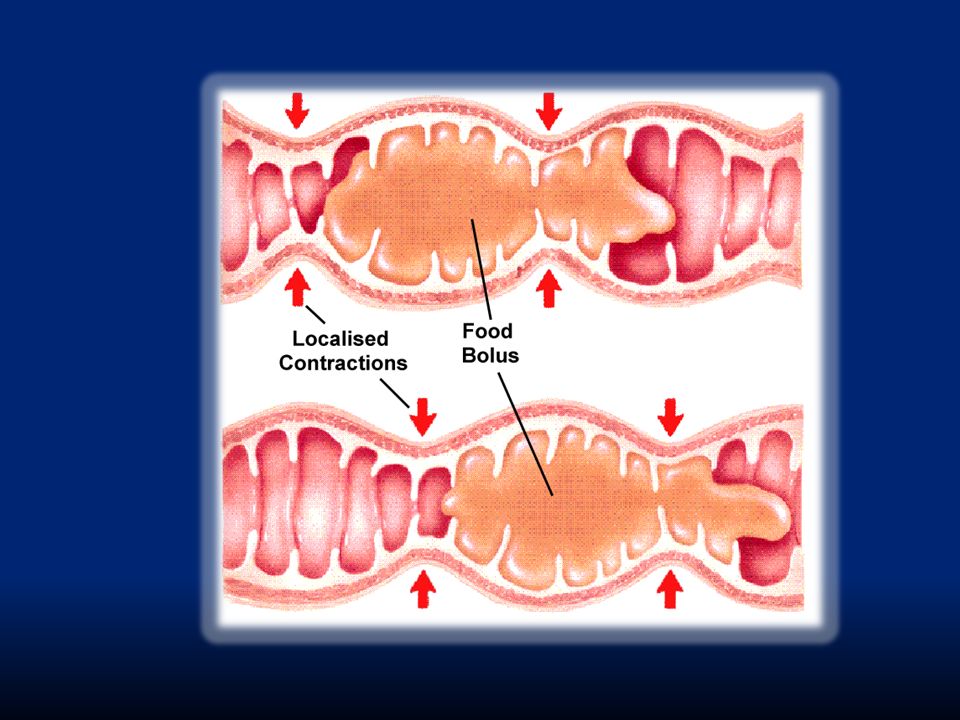
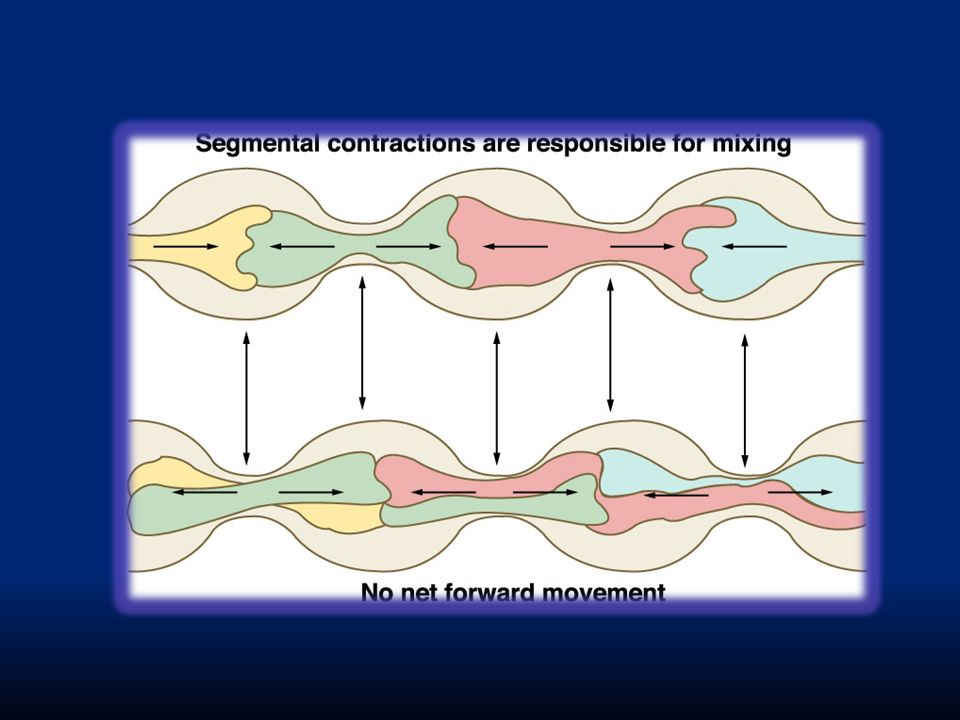
B. Questa motilità include: i. Movimenti di segmentazione per ottimizzare l’assorbimento: si generano contrazioni concentriche localizzate e spaziate lungo l’intestino. La lunghezza di ogni contrazione è circa 1cm. Sono determinate dalle cellule di Cajal ii. Movimenti peristaltici per far avanzare il contenuto iii. Movimenti di massa per svuotare rapidamente il contenuto luminale

34
i.L’aspetto è quello di una catena di salsicce. Quando finisce un’ondata di queste contrazioni, ne inizia un’altra con “salsicciotti” intermedi ai precedenti. In questo modo il cibo viene frammentato molto finemente. La frequenza di contrazione è di 8-12 contrazioni al minuto per duodeno e digiuno.

37
ii.Le onde peristaltiche determinano una velocità di progressione di 0.5-2 cm/s. Si estinguono dopo 3-5 cm. Il movimento del cibo nell’intestino tenue è quindi molto lento e vi staziona per circa 3-5 ore. La peristalsi aumenta moltissimo dopo l’ingestione di un pasto per circuiti dovuti alla distensione dell’organo. La distensione dello stomaco per l’arrivo di cibo innesca un’attivazione del plesso mienterico che si propaga fino all’intestino.

38
Inoltre vari ormoni innescano questi movimenti: Gastrina CCK Secretina Insulina Glucagone Serotonina Stimolano la peristalsi peristalsi Inibiscono la peristalsi peristalsi

39
La funzione della peristalsi a questo livello non è soltanto quella di spingere il chimo verso la valvola ileocecale, ma anche quello di far aderire il cibo verso le pareti dell’intestino in modo da facilitare l’ingresso nel duodeno di altro cibo. In genere fino a quando non viene ingerito altro cibo la valvola ileocecale resta chiusa. Solo un nuovo riflesso gastroenterico la fa aprire e permette il passaggio del chimo nell’intestino cieco.

41
iii.I movimenti di massa sono dovuti a contrazioni sostenute e di lunga durata della muscolatura circolare. Interessano lunghi tratti e si ripetono ogni 2-3 ore. Rispetto ai precedenti questi movimenti hanno una forza di contrazione maggiore, un tempo prolungato di manifestazione e sono bidirezionali (verso l’orifizio anale e verso l’orifizio orale). Nell’Uomo, in condizioni fisiologiche, sono assenti nell’intestino tenue, ma possono comparire in seguito a tossicosi alimentari, a fattori patologici del canale alimentare.
. Nell’Uomo, in condizioni fisiologiche, sono assenti nell’intestino tenue, ma possono comparire in seguito a tossicosi alimentari, a fattori patologici del canale alimentare..")
42
Funzioni della valvola ileo-cecale La funzione principale è quella di prevenire il reflusso di cibo dal colon all’intestino tenue. I lembi della valvola protrudono nel lume del cieco e restano quindi chiusi quando il chimo tende a refluire verso l’ileo. Inoltre la valvola è fornita di uno sfintere che la mantiene chiusa rallentando lo svuotamento dell’ileo. Subito dopo il pasto il riflesso gastro-ileale intensifica la peristalsi nell’ileo. Ogni giorno passano attraverso la valvola circa 1500 ml di chimo.

43
Lo stato di contrazione dello sfintere ileocecale e l’intensità della peristalsi sono anche sotto il controllo dell’intestino cieco. Quando questo è disteso lo sfintere viene mantenuto chiuso e la peristalsi notevolmente ridotta. Questi riflessi sono mediati soprattutto dal SNE.

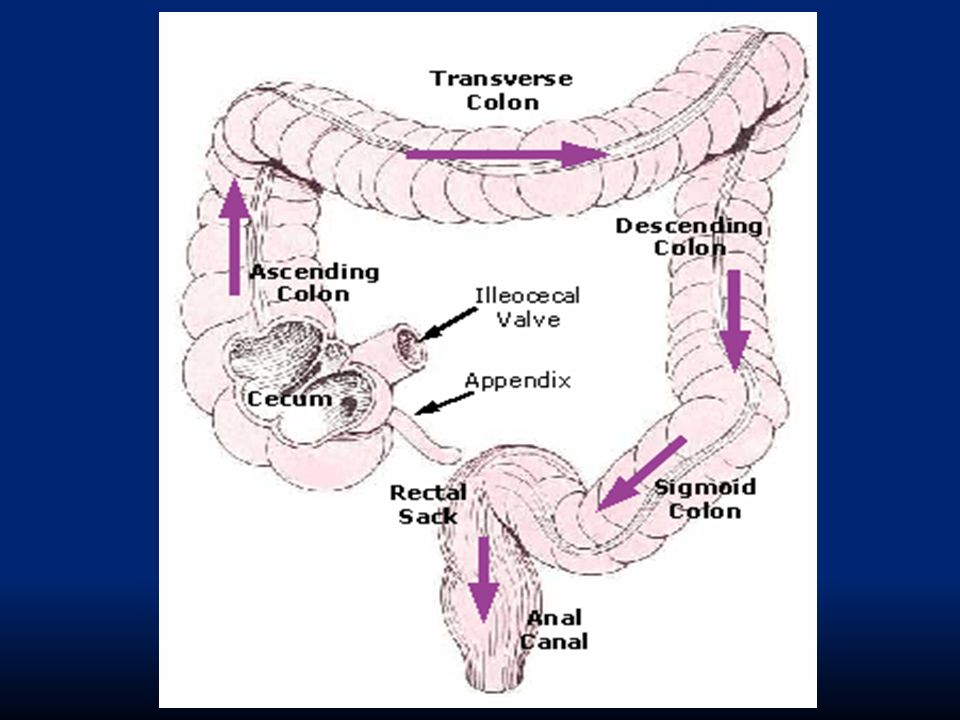
44
Le funzioni motorie principali del cieco-colon sono: Miscelare il materiale per favorire l’assorbimento di acqua ed elettroliti Generare un’attività propulsiva che assicuri il transito intestinale Immagazzinamento temporaneo di materia fecale e attuazione dei processi evacuativi

46
Si trovano i seguenti tipi di movimento: Segmentazione: contrazioni poco organizzate, con il solo scopo di aumentare la pressione per spingere il materiale. Dette anche haustrazioni. Dovute alla caratteristica distribuzione della muscolatura longitudinale che si organizza in tre fasci sottili detti tenie. Peristalsi: dopo il pasto Movimenti di massa: colon trasverso e discendente

47
Immagine radiografica del colon di un individuo normale in cui sono evidenti la distribuzione e la configurazione delle haustre

48
In particolare la prima parte del colon è destinata all’assorbimento mentre la seconda parte allo storage. Il colon: riceve dall’ileo 1500 ml di chilo/die riassorbe il 95% di H 2 O e sali 1.2-1.7 m di lunghezza bassa velocità di propulsione (5-10 cm/ora) il chilo rimane 18-24 ore nel colon
 il chilo rimane ore nel colon.")
49
L’intestino retto è solitamente vuoto. Quando il materiale spinto dal movimento di massa arriva nel retto, ha inizio lo stimolo della defecazione. Il materiale è trattenuto grazie a: Sfintere anale interno: anello di muscolatura liscia subito interno all’ano (involontario) Sfintere anale esterno: formato da muscolatura striata. Controllato dai nervi pudenda.
 Sfintere anale esterno: formato da muscolatura striata. Controllato dai nervi pudenda..")
50
Quando il retto si riempie, la distensione della parete inizia un riflesso mienterico con aumento della peristalsi in colon, sigma e retto che forza il materiale verso l’ano rilasciando lo sfintere interno. In realtà il meccanismo è rafforzato da un feed-back che utilizza i nervi pelvici e fa capo al sistema parasimpatico.

52
SecrezioneAssorbimento Ghiandole salivari 1.5 l/die --- Stomaco 2.0 l/die minimo Succo pancreatico +bile 3.0 l/die --- Intestino tenue 1.5 l/die 7.0 l/die Colon--- 0.7 l/die Totale 8 l/die 7.7 l/die Perdita con le feci 300 ml/die

53
Nel tratto gi le ghiandole secretorie hanno due funzioni fondamentali: secernono enzimi digestivi lungo tutto il tratto, dalla bocca all’intestino secernono muco a funzione lubrificante In generale le secrezioni digestive avvengono solo in presenza di cibo e sono differenti a seconda del tipo di cibo presente. Vediamo i vari tipi di secrezione e prima qualche principio generale per quanto riguarda la funzione secretoria nel tratto digerente.

54
Esistono vari tipi di ghiandole: Cellule mucose: sono cellule singole che secernono muco. Sono attivate dall’irritazione dell’epitelio circostante e producono muco che agisce come lubrificante e protegge la superficie epiteliale da escoriazioni e succhi digestivi. Pozzetti ghiandolari: sono tratti di epitelio ricchi di invaginazioni profonde che contengono cellule secernenti (ad esempio le cripte di Lieberkühn)
.")
55
Ghiandole tubulari: vedi ghiandole gastriche e duodenali Complessi ghiandolari: ghiandole salivari, pancreas e fegato. Le altre ghiandole sono formate da acini.

56
La secrezione delle ghiandole del tratto gi è stimolato soprattutto dal contatto con il cibo. Inoltre la stimolazione epiteliale attiva il SNE attraverso: stimolazione tattilestimolazione tattile irritazione chimicairritazione chimica distensione della paretedistensione della parete I neuroni del plesso sottomucoso stimolano la secrezione ghiandolare.

57
STIMOLAZIONE PARASIMPATICA: determina forte stimolazione nella secrezione delle ghiandole salivari, esofagee, gastriche, del pancreas e duodenali STIMOLAZIONE SIMPATICA: può debolmente stimolare l’attività secretoria, ma poiché determina vasocostrizione dei vasi che irrorano la ghiandola, molto spesso la stimolazione simpatica inibisce la secrezione per ridotto afflusso sanguigno

58
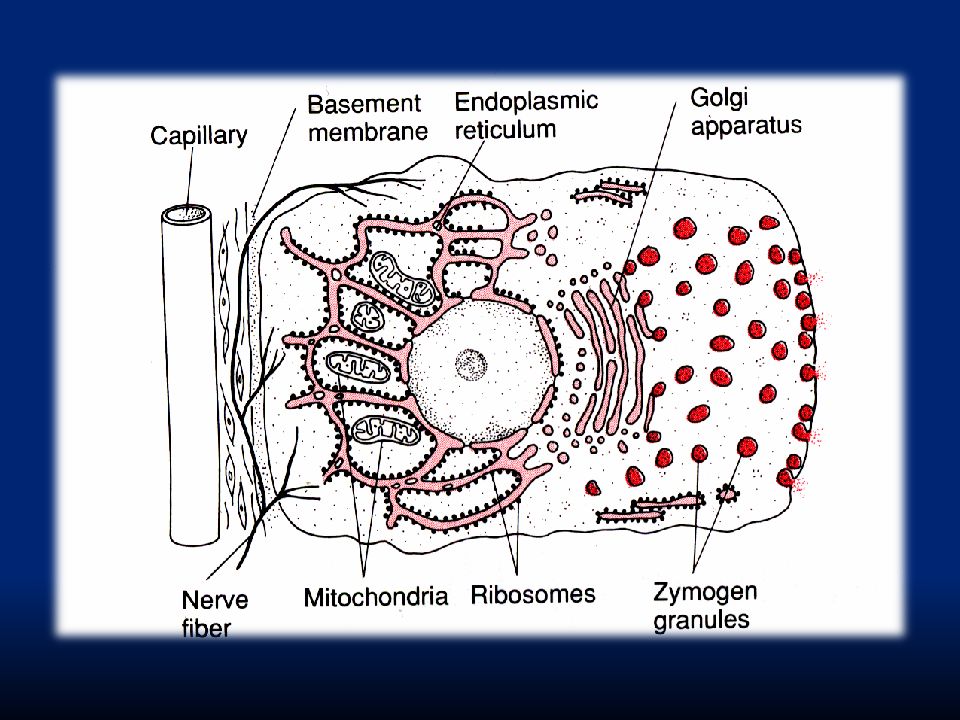
Secrezione delle cellule ghiandolari: i. materiale precursore trasportato per diffusione o trasporto attivo alla base della cellula ghiandolare ii. formazione di ATP dai mitocondri iii. sintesi di sostanze organiche nel reticolo endoplasmico e nel Golgi iv. trasporto del materiale secretorio v. formazione delle vescicole secretorie nel complesso del Golgi vi. esocitosi Ca-dipendente delle vescicole secretorie

59
capillare membrana basale con ingresso di sostanze secrezione dei granuli di zimogeno

61
Immagine di una cellula ghiandolare basale con reticolo endoplasmico apicale con granuli di zimogeno

62
Il muco è composto da acqua, elettroliti, glicoproteine. È leggermente differente nelle varie porzioni del tratto gi, ma ha comunque le stesse funzioni: proprietà adesive che fanno aderire il cibo alle pareti dell’organo protegge la mucosa dall’azione degli enzimi digestivi azione lubrificante per far scorrere il cibo forma la massa fecale facendo aderire le particelle le glicoproteine svolgono un’azione protettiva contro variazioni del pH (in genere è presente bicarbonato)
.")
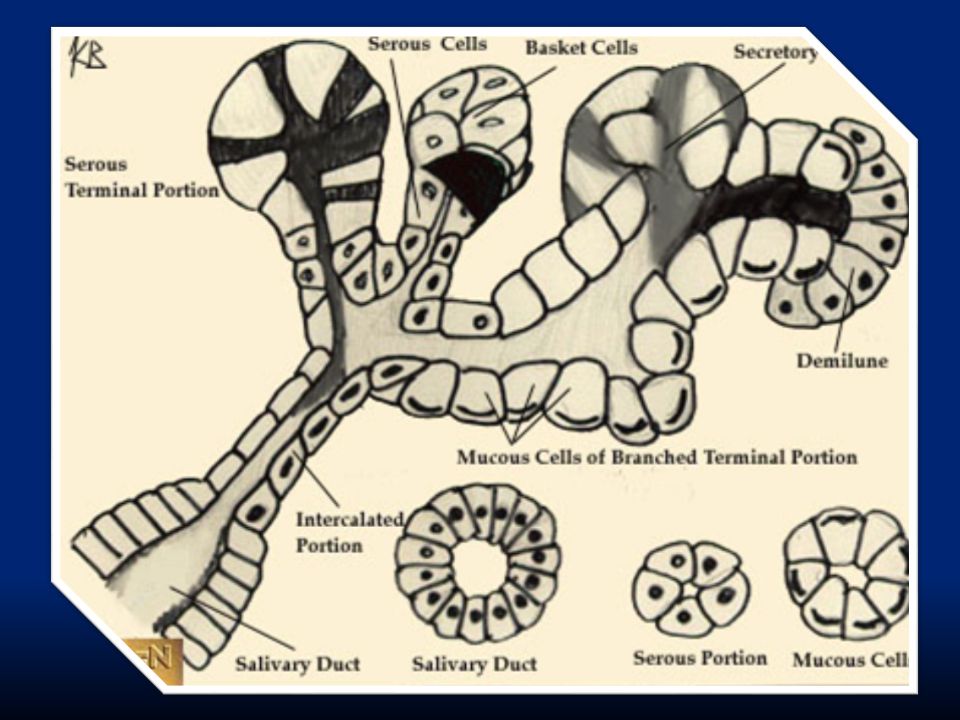
63
La secrezione avviene ad opera delle ghiandole salivari (ghiandole specializzate formate da acini e dotti salivari) che producono 1000-1500ml/die: PAROTIDI: secrezione sierosa priva di mucina SOTTOMANDIBOLARI: secrezione mista sierosa e mucosa SOTTOLINGUALI: secrezione mista sierosa e mucosa In più ci sono molte ghiandole buccali che secernono solo muco.
 che producono ml/die: PAROTIDI: secrezione sierosa priva di mucina SOTTOMANDIBOLARI: secrezione mista sierosa e mucosa SOTTOLINGUALI: secrezione mista sierosa e mucosa In più ci sono molte ghiandole buccali che secernono solo muco.")
64
Distribuzione della saliva a riposo

65
Dotto di Stenone (parotide) Dotto di Wharton (sottomandibolare) Dotto di Bartolino (sottolinguale)
 Dotto di Wharton (sottomandibolare) Dotto di Bartolino (sottolinguale)")
67
La saliva ha diverse funzioni: Lubrifica il cibo Facilita la parola Diluisce le sostanze nocive Inizia la digestione dell’amido Neutralizza gli acidi Ha un’azione protettiva sui denti È importante per la sensazione di sete

68
La saliva contiene diverse sostanze organiche. I composti organici sono costituiti prevalentemente da proteine e glicoproteine. La ptialina (amilasi salivare) (digerisce l’amilosio) e la mucina (a funzione lubrificante) costituiscono la maggior parte della secrezione organica. La saliva contiene anche una certa quota di albumina, immunoglobuline, tiocianato ed altre sostanze ad azione batteriostatica o addirittura battericida, per contrastare la proliferazione batterica, quali sialoperossidasi e lisozima.
 (digerisce l’amilosio) e la mucina (a funzione lubrificante) costituiscono la maggior parte della secrezione organica. La saliva contiene anche una certa quota di albumina, immunoglobuline, tiocianato ed altre sostanze ad azione batteriostatica o addirittura battericida, per contrastare la proliferazione batterica, quali sialoperossidasi e lisozima..")
69
α-amilasi idrolizza i legami α -1,4 dell’amido (componente detta amilosio), ma non ha effetto sui legami α-1,6 (componente detta amilopectina) responsabili dell’elevato grado di ramificazione.
, ma non ha effetto sui legami α-1,6 (componente detta amilopectina) responsabili dell’elevato grado di ramificazione.")
70
I due tipi di secrezione proteica sono costituiti da: Secrezione sierosa che contiene ptialina (un’isoforma dell’ α-amilasi): è un enzima che serve per la digestione dell’amido (l'organismo umano non dispone di alcun enzima per la cellulosa). Il tempo di azione della ptialina é ridotto, ma supera quello della permanenza del bolo nella cavità orale in quanto prosegue nello stomaco, fino all’inattivazione per denaturazione operata dall'acido cloridrico. Secrezione mucosa che contiene mucina per azione protettiva e lubrificante.

71
La saliva ha un pH compreso fra 6 e 7. Il flusso è variabile fra 0.1 ml/min per aumentare fino a 4 ml/min. Composizione ionica, osmolarità e pH variano con il flusso. La saliva contiene grandi quantità di bicarbonato e potassio. Sodio e cloro sono molto meno concentrati che nel plasma.

72
Apertura delle ghiandole salivari nella cavità buccale

73
La secrezione salivare avviene in due steps: I. Gli acini secernono una saliva primaria che contiene ptialina e/o mucina in una soluzione con una concentrazione ionica simile a quella del LEC. Questa comincia a fluire nei dotti e qui avvengono due processi.

74
II. Lungo i dotti (impermeabili all’acqua) avviene una modificazione legata al riassorbimento attivo di Na + e alla secrezione di K +. Cl - è passivamente riassorbito e scambiato con HCO 3 -. La saliva finale è ipotonica rispetto al plasma
 avviene una modificazione legata al riassorbimento attivo di Na + e alla secrezione di K +. Cl - è passivamente riassorbito e scambiato con HCO 3 -. La saliva finale è ipotonica rispetto al plasma.")
75
Quando il flusso salivare aumenta: Aumentano le concentrazioniAumentano le concentrazioni di NaCl perché Na + e Cl - sono riassorbiti dall’epitelio dei dotti più lentamente Aumenta la permeabilitàAumenta la permeabilitàall’acqua Esiste una secrezione continua di HCO 3 -Esiste una secrezione continua di HCO 3 - Come risultato l’osmolarità tende ad aumentare e il pH diventa basico

76
Quindi in condizioni di riposo si ha: [Na + ] e [Cl - ] circa 15mEq/l [K + ] circa 30mEq/l [HCO 3 - ] circa 50-70mEq/l A differenza delle altre secrezioni ghiandolari gastrointestinali, la secrezione salivare non è sotto controllo ormonale, ma piuttosto sotto totale controllo nervoso da parte del sistema autonomo.
![Quindi in condizioni di riposo si ha: [Na + ] e [Cl - ] circa 15mEq/l [K + ] circa 30mEq/l [HCO 3 - ] circa 50-70mEq/l A differenza delle altre secrezioni ghiandolari gastrointestinali, la secrezione salivare non è sotto controllo ormonale, ma piuttosto sotto totale controllo nervoso da parte del sistema autonomo.](http://images.slideplayer.it/33/10192317/slides/slide_76.jpg "Quindi in condizioni di riposo si ha: [Na + ] e [Cl - ] circa 15mEq/l [K + ] circa 30mEq/l [HCO 3 - ] circa 50-70mEq/l A differenza delle altre secrezioni ghiandolari gastrointestinali, la secrezione salivare non è sotto controllo ormonale, ma piuttosto sotto totale controllo nervoso da parte del sistema autonomo.")
77
Le ghiandole salivari sono innervate da ramificazioni del sistema parasimpatico che prendono origine dai nuclei salivatori superiore (VII) ed inferiore (IX) del tronco encefalico. Gli stimoli per l’attivazione di questi centri sono: stimoli gustativi stimoli gustativi stimoli tattili provenienti dalla cavità orale stimoli tattili provenienti dalla cavità orale stimoli provenienti da centri nervosi superiori legati all’odore o alla consistenza del cibo stimoli provenienti da centri nervosi superiori legati all’odore o alla consistenza del cibo stimoli gastrici o intestinali stimoli gastrici o intestinali

78
Fb dal n. salivatorio sup. con fb postgangliari a livello di ganglio sottolinguale e sottomandibolare Fb dal n. salivatorio inf. con fb postgangliari a livello di ganglio otico

79
Il riflesso salivare primario è innato, cioè INCONDIZIONATO. Tuttavia la secrezione salivare può essere attivata anche da RIFLESSI CONDIZIONATI CENTRALI, acquisiti con l’esperienza (cane di Pavlov)
.")
80
Pavlov stava studiando le secrezioni della ghiandola salivare dei cani quando notò che esse iniziavano a secernere PRIMA del contatto con il cibo. È noto il suo classico esperimento: associando per un certo numero di volte la presentazione della carne al suono di un campanello, alla fine il solo suono del campanello determina la salivazione nel cane, che prima certamente non produceva; quindi il cane si comporta così a causa di un riflesso condizionato provocato in lui artificialmente

81
Dalla cavità orale il bolo passa nell’esofago (dove staziona per circa 10 secondi) che funziona come via di comunicazione con lo stomaco. L’esofago sfocia nello stomaco a livello dello sfintere esofageo inferiore o sfintere gastro-esofageo (cardias). Le secrezioni esofagee sono interamente mucose a funzione lubrificante. La secrezione avviene attraverso ghiandole mucose.
. Le secrezioni esofagee sono interamente mucose a funzione lubrificante. La secrezione avviene attraverso ghiandole mucose..")
82
Esistono cellule muco-secernenti sempre a funzione protettiva e lubrificante e ghiandole tubulari che sono: Ghiandole ossintiche o gastriche: superficie interna del corpo e del fondo. Secernono pepsinogeno (cellule peptiche o principali), HCl e fattore intrinseco (cellule ossintiche o parietali). Ghiandole piloriche: sono nell’antro e secernono muco protettivo, pepsinogeno e gastrina.
, HCl e fattore intrinseco (cellule ossintiche o parietali). Ghiandole piloriche: sono nell’antro e secernono muco protettivo, pepsinogeno e gastrina..")
84
Cellule mucosali del collo Cellule peptiche o principali che secernono pepsinogeno Cellule parietali o ossintiche che secernono HCl e fattore intrinseco

86
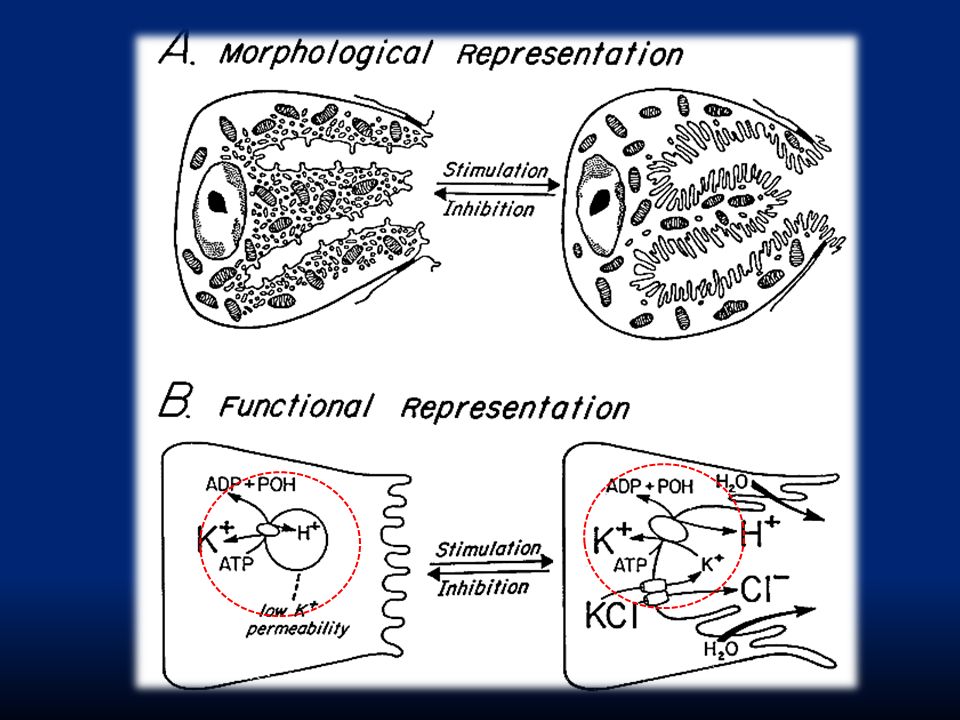
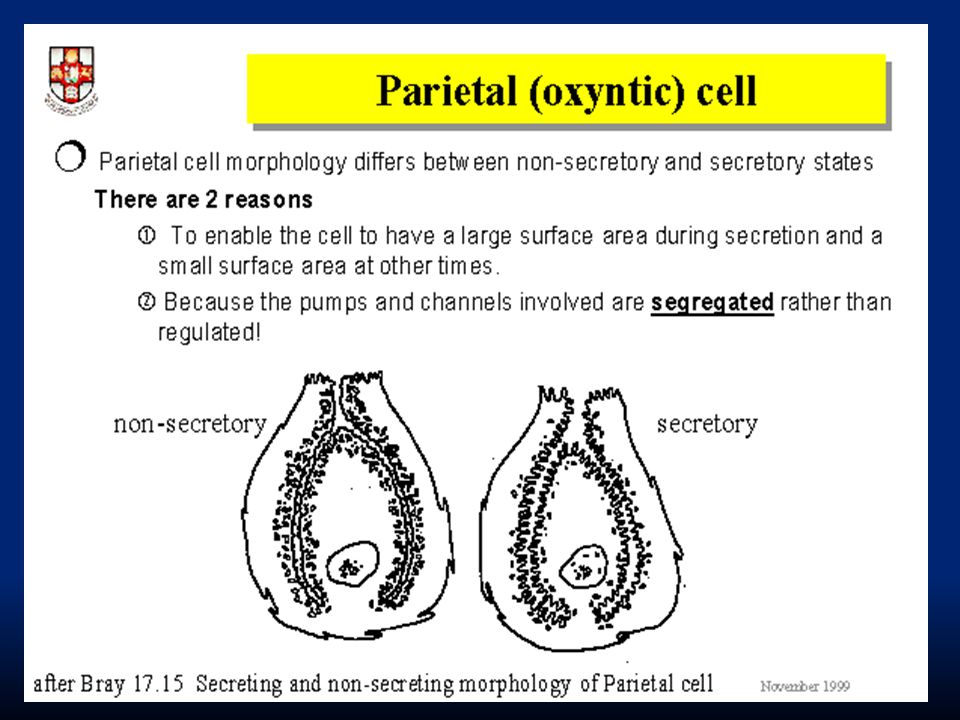
LUME Caratteristica modificazione morfo-funzionale delle cellule parietali secernenti. HCl è prodotto a livello dei canalicoli intracellulari e viene poi riversato nel lume dello stomaco

87
La concentrazione di H + nelle secrezioni delle cellule parietali è circa 3·10 6 volte superiore che nel sangue e Cl - è secreto contro un gradiente elettrico e chimico. Quindi, la capacità delle cellule parietali di secernere HCl dipende da meccanismi di trasporto attivo. Il ruolo chiave è la pompa H + /K + ATPasi localizzata nella membrana delle strutture tubulo-vescicolari.

89
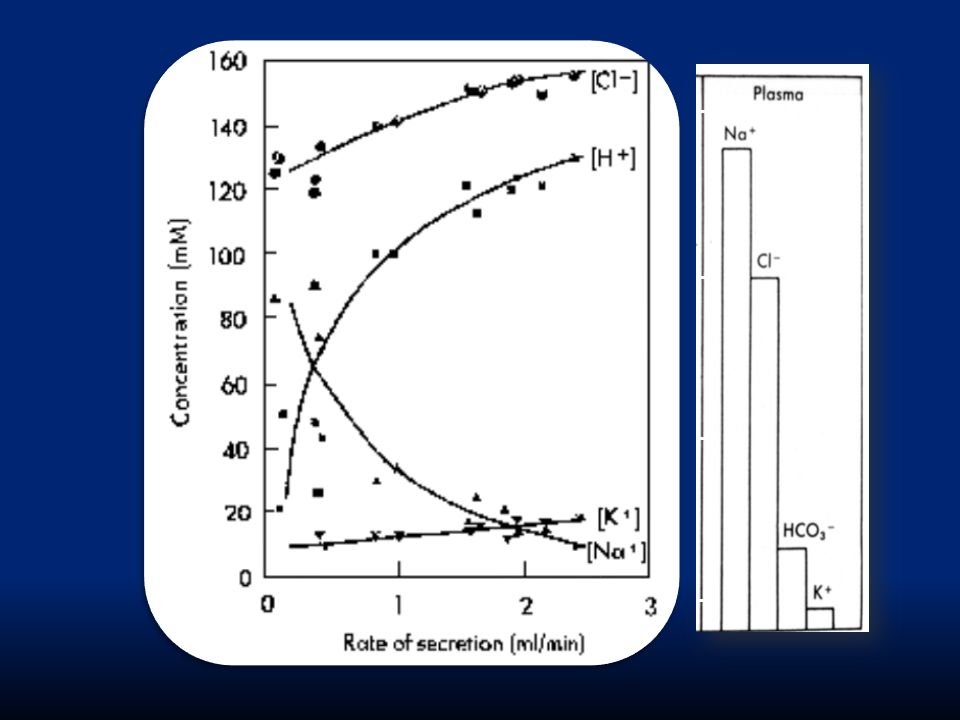
1.La reazione di idratazione di CO 2, che proviene dal metabolismo cellulare e dall’esterno, ad opera della AC forma acido carbonico H 2 CO 3 che si dissocia subito in H + e HCO3 -. Gli idrogenioni sono pompati nel lume da una H + /K + ATPasi. 2. HCO 3 - è scambiato con Cl - (montata alcalina). 3.Cl - e K + sono trasportati nel lume; il K + quindi ha effettivamente un riciclo 4.L’accumulo di ioni H + osmoticamente attivi genera un gradiente osmotico attraverso la membrana che risulta in diffusione di acqua verso l’esterno con formazione di un succo gastrico contenente 155 mM HCl e 15 mM KCl.
. 3.Cl - e K + sono trasportati nel lume; il K + quindi ha effettivamente un riciclo 4.L’accumulo di ioni H + osmoticamente attivi genera un gradiente osmotico attraverso la membrana che risulta in diffusione di acqua verso l’esterno con formazione di un succo gastrico contenente 155 mM HCl e 15 mM KCl..")
90
1. La reazione di idratazione di CO 2, che proviene dal metabolismo cellulare e dall’esterno, ad opera della AC forma acido carbonico H 2 CO 3 che si dissocia subito in H + e HCO 3 -. Gli idrogenioni sono pompati nel lume da una H + /K + ATPasi. 2. HCO 3 - è scambiato con Cl - (montata alcalina). 3. K + è trasportato nel lume iperpolarizzando la cellula e richiamando Cl -. 4. L’accumulo di ioni H + osmoticamente attivi genera un gradiente osmotico attraverso la membrana che risulta in diffusione di acqua verso l’esterno con formazione di un succo gastrico contenente 155 mM HCl e 15 mM KCl.
. 3. K + è trasportato nel lume iperpolarizzando la cellula e richiamando Cl L’accumulo di ioni H + osmoticamente attivi genera un gradiente osmotico attraverso la membrana che risulta in diffusione di acqua verso l’esterno con formazione di un succo gastrico contenente 155 mM HCl e 15 mM KCl..")
91
Na + e K + restano quindi poco concentrati nel lume, mentre si forma una soluzione ricca di HCl con 150mEq/l di HCl. L’azione dell’enzima anidrasi carbonica è fondamentale, tanto che se questa è inibita da acetazolamide, la secrezione di HCl non avviene.

93
G = gastrina, A = ACh, H = istamina; G = gastrina, A = ACh, H = istamina; la H + /K + -ATPasi si attiva quando, in seguito a stimoli secretagoghi, si ha la fusione degli organuli tubulo- vescicolari (T) con la membrana plasmatica e la formazione dei canalicoli secretori (cs); a seguito della sua traslocazione su queste invaginazioni della membrana esterna, la pompa può legare K + la H + /K + -ATPasi si attiva quando, in seguito a stimoli secretagoghi, si ha la fusione degli organuli tubulo- vescicolari (T) con la membrana plasmatica e la formazione dei canalicoli secretori (cs); a seguito della sua traslocazione su queste invaginazioni della membrana esterna, la pompa può legare K +
 con la membrana plasmatica e la formazione dei canalicoli secretori (cs); a seguito della sua traslocazione su queste invaginazioni della membrana esterna, la pompa può legare K + la H + /K + -ATPasi si attiva quando, in seguito a stimoli secretagoghi, si ha la fusione degli organuli tubulo- vescicolari (T) con la membrana plasmatica e la formazione dei canalicoli secretori (cs); a seguito della sua traslocazione su queste invaginazioni della membrana esterna, la pompa può legare K +")
95
Muco e bicarbonato: funzione protettiva dall’ambiente acido dello stomaco per presenza del bicarbonato. Formano uno strato che può essere anche di 1 cm. È facilmente attaccabile da composti quali anitinfiammatori e aspirina. Fattore intrinseco di Castle: serve per l’assorbimento a livello intestinale della vitamina B 12. La sua mancanza causa anemia perniciosa (macrocitica) per mancata maturazione dei globuli rossi. Il fattore intrinseco si lega alla cianocobalamina impedendo che la flora batterica la possa utilizzare
 per mancata maturazione dei globuli rossi. Il fattore intrinseco si lega alla cianocobalamina impedendo che la flora batterica la possa utilizzare.")
96
Funzione del fattore intrinseco (IF) nell’assorbimento della vitamina B 12 a livello ileale
 nell’assorbimento della vitamina B 12 a livello ileale")
97
Pepsinogeno*: precursori (ne esistono diversi) delle pepsine che sono proteasi attive a pH acidi. Si attivano quando vengono in contatto con HCl. La migliore attività si ha per pH ≤ 3. Si neutralizzano appena il pH si sposta a valori più alti. Sono secrete anche piccole quantità di altri enzimi fra cui la lipasi gastrica, l’amilasi gastrica e la gelatinasi per i proteoglicani della carne. Sono poco attivi. HCl: serve a creare l’ambiente acido per l’attivazione dei pepsinogeni. *Le pepsine riconoscono aa aromatici ma non gli altri, quindi iniziano soltanto il processo di digestione delle proteine. Esistono almeno tre diversi tipi di pepsine: Pepsina I: pH ottimale di 3-3.2Pepsina I: pH ottimale di 3-3.2 Pepsina II e III. Secrete solo nel corpo dello stomaco hanno pH ottimale di 1.5-2Pepsina II e III. Secrete solo nel corpo dello stomaco hanno pH ottimale di 1.5-2

98
Il muco ha funzione protettiva perché forma una barriera che impedisce all’acido cloridrico di danneggiare la mucosa

99
L’ulcera gastrica è una grave patologia di cui è responsabile nella maggior parte dei casi l’infezione da Helicobacter pylori (che in casi meno gravi determina gastrite) che attacca la mucosa gastrica distruggendola e provocando la morte delle cellule mucosali.
 che attacca la mucosa gastrica distruggendola e provocando la morte delle cellule mucosali.")
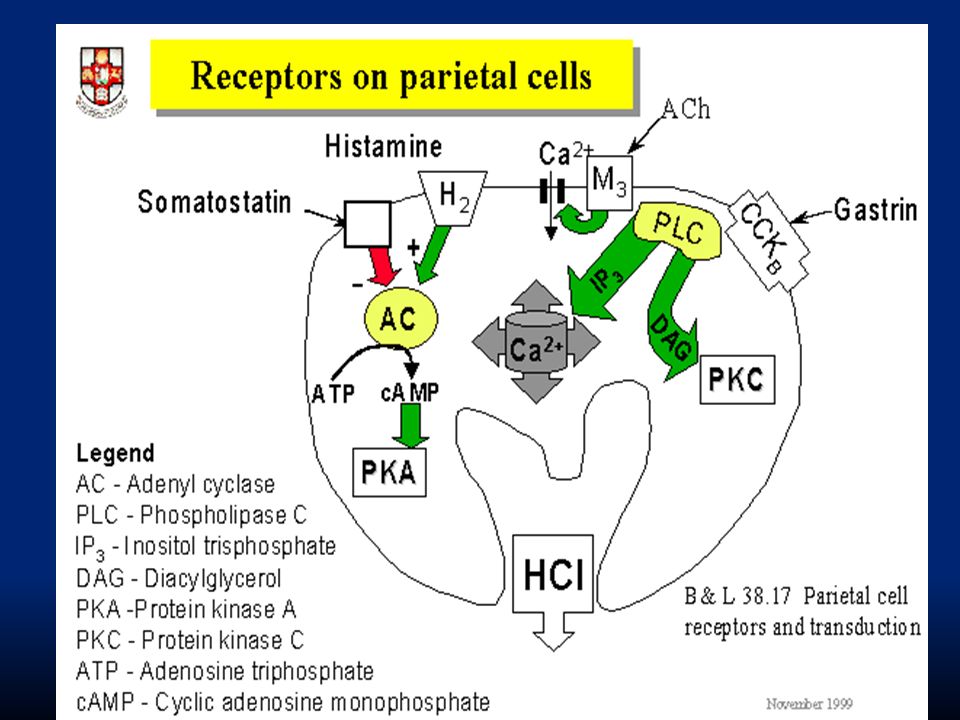
100
I fattori principali per la secrezione di HCl sono: ACh: rilasciata dai terminali colinergici vagali. Si lega ai recettori M3. Ha azione su tutti i tipi cellulari (principali, parietali e mucosali) Istamina: proviene dalle cellule della mucosa gastrica. Si lega ai recettori H2. Ha effetto sulla secrezione di HCl. Gastrina: proviene dalle cellule G della mucosa dell’antro. Ha effetto sulla secrezione di HCl. Attraverso meccanismi che aumentano il calcio intracellulare inducendo esocitosi
 Istamina: proviene dalle cellule della mucosa gastrica. Si lega ai recettori H2. Ha effetto sulla secrezione di HCl. Gastrina: proviene dalle cellule G della mucosa dell’antro. Ha effetto sulla secrezione di HCl. Attraverso meccanismi che aumentano il calcio intracellulare inducendo esocitosi.")
102
Circa la metà dei segnali che causano secrezione gastrica originano nel nucleo motorio dorsale del vago e decorrono nel vago fino ai plessi del SNE. L’altra metà ha origine direttamente nel SNE. Gli stimoli che danno inizio al processo di secrezione sono: i. distensione della parete gastrica ii. stimolazione tattile della mucosa iii. stimolazione chimica ad opera di aa e peptidi

103
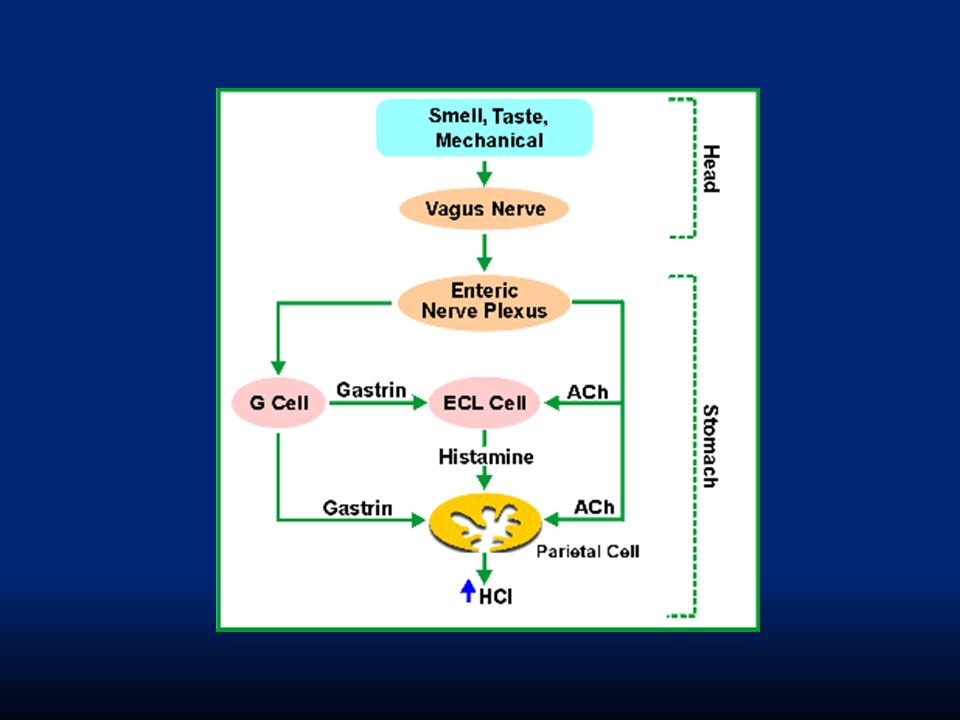
ACh: stimoli opportuni quali la distensione dello stomaco, stimoli tattili sulla mucosa e stimoli chimici (alcuni aa e peptidi) iniziano una serie di riflessi nervosi che portano al rilascio di ACh dai terminali vagali sulle cellule delle ghiandole gastriche stimolando la secrezione. Il rilascio di ACh è indotto già durante la fase cefalica da stimoli tattili della cavità orale e da fattori centrali quali vista, odore o ricordo del cibo

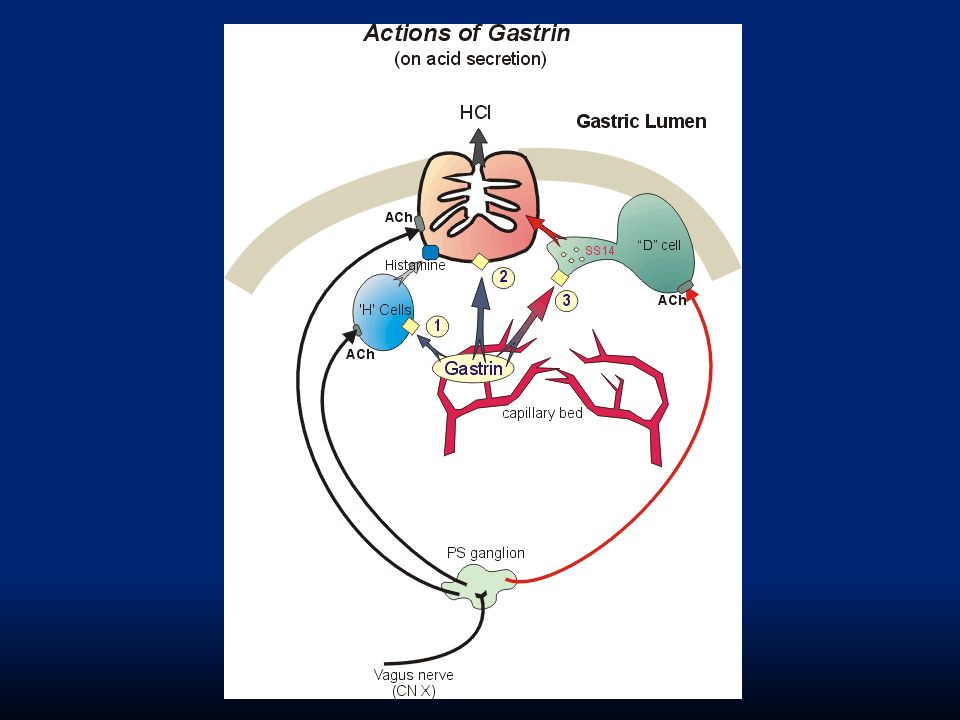
104
Gastrina: la stimolazione nervosa delle cellule presenti nell’antro e secernenti gastrina (G cells) induce il rilascio dell’ormone. Questo viene veicolato dal torrente sanguigno e arriva alle cellule principali e peptiche stimolandone la secrezione.

105
: è continuamente formata nella mucosa gastrica dalle cellule enterocromaffini. È il più potente secretagogo fisiologico. Da sola è poco efficace nello stimolare la secrezione gastrica, ma è un fondamentale co-fattore per l’azione della gastrina e di ACh. Se viene inibita l’azione dell’istamina con cimetidina, anche l’azione di ACh e gastrina è molto ridotta. Istamina: è continuamente formata nella mucosa gastrica dalle cellule enterocromaffini. È il più potente secretagogo fisiologico. Da sola è poco efficace nello stimolare la secrezione gastrica, ma è un fondamentale co-fattore per l’azione della gastrina e di ACh. Se viene inibita l’azione dell’istamina con cimetidina, anche l’azione di ACh e gastrina è molto ridotta.

106
I tre fattori ACh, istamina e gastrina determinano un forte sinergismo che potenzia moltissimo la secrezione di HCl

109
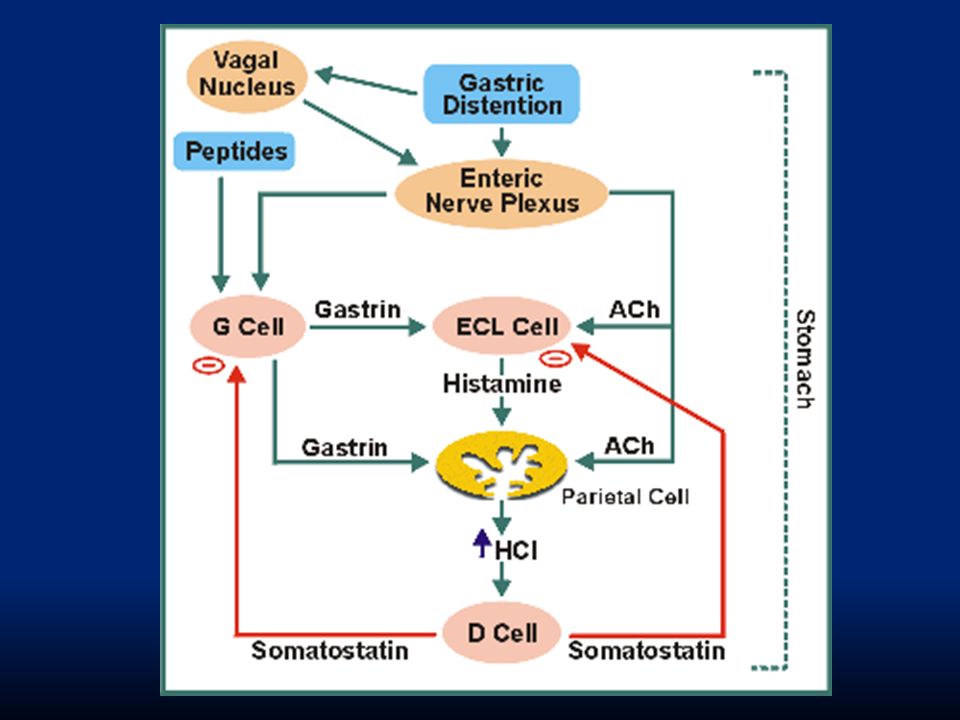
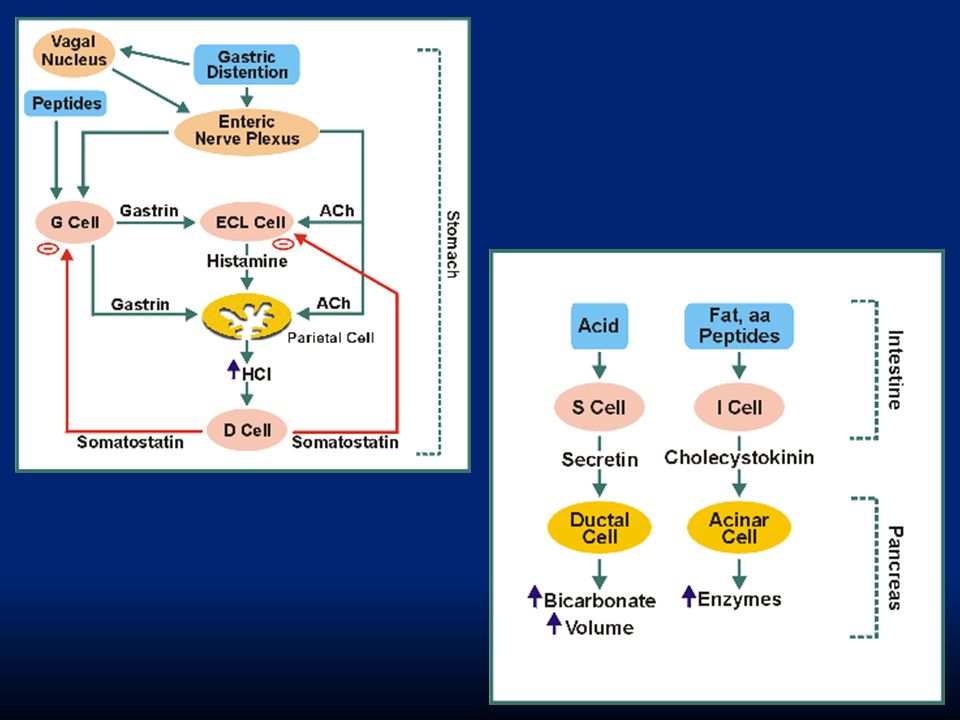
La regolazione del rilascio di pepsinogeno è più semplice. Avviene in risposta a: i. stimolazione delle cellule principali con ACh rilasciata dal vago o dai neuroni enterici ii. stimolazione delle cellule in risposta all’ambiente acido Va ricordato che quando l’acidità dello stomaco aumenta troppo interviene un feed-back di controllo ad opera della somatostatina che inibisce il rilascio di gastrina dalle cellule G e impedisce ulteriore rilascio di HCl.

110
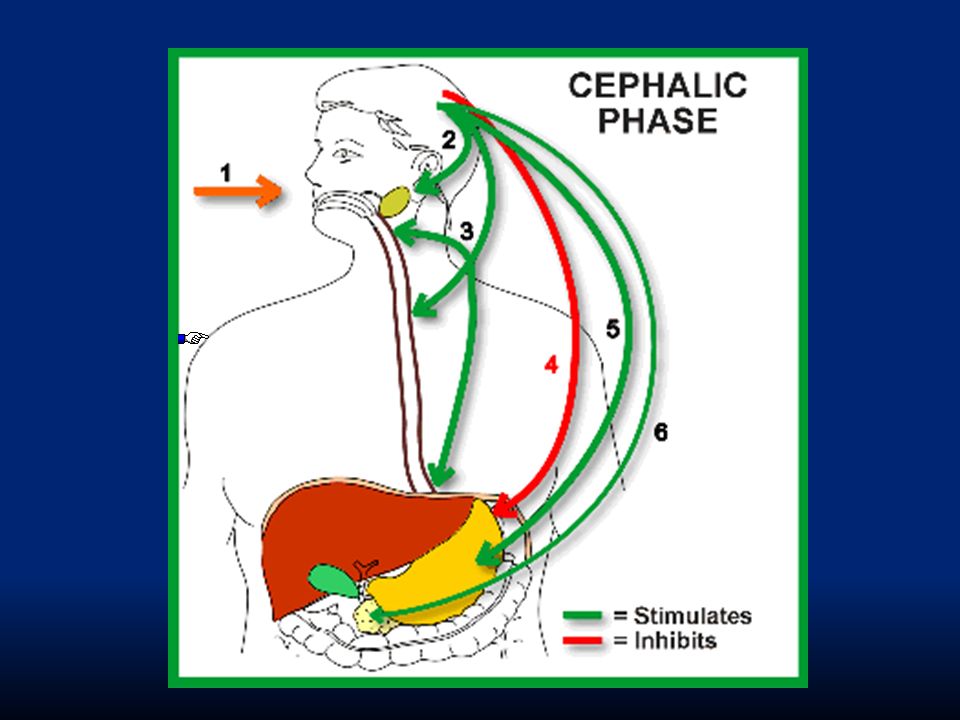
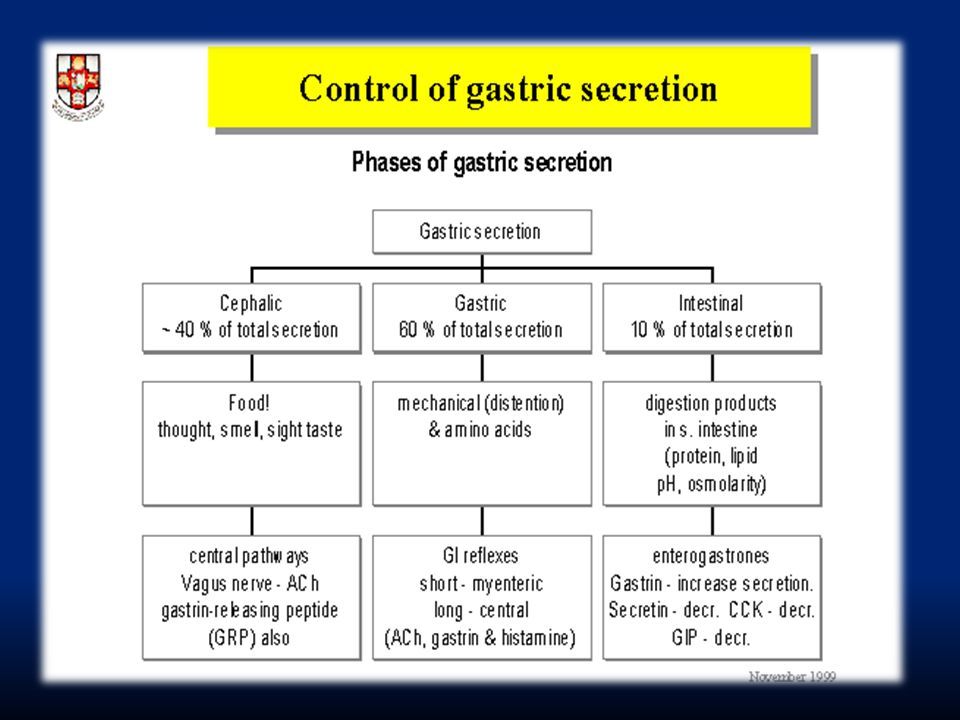
Fase cefalica: rende conto del 20% della secrezione gastrica. È evocata dalla vista, dal ricordo e dall’idea del cibo. Avviene anche prima che il cibo arrivi allo stomaco (pasto fittizio di Pavlov). I segnali nervosi originano dalla corteccia o nei centri dell’appetito e sono inviati allo stomaco attraverso il nucleo motorio dorsale del nervo vago.
. I segnali nervosi originano dalla corteccia o nei centri dell’appetito e sono inviati allo stomaco attraverso il nucleo motorio dorsale del nervo vago..")
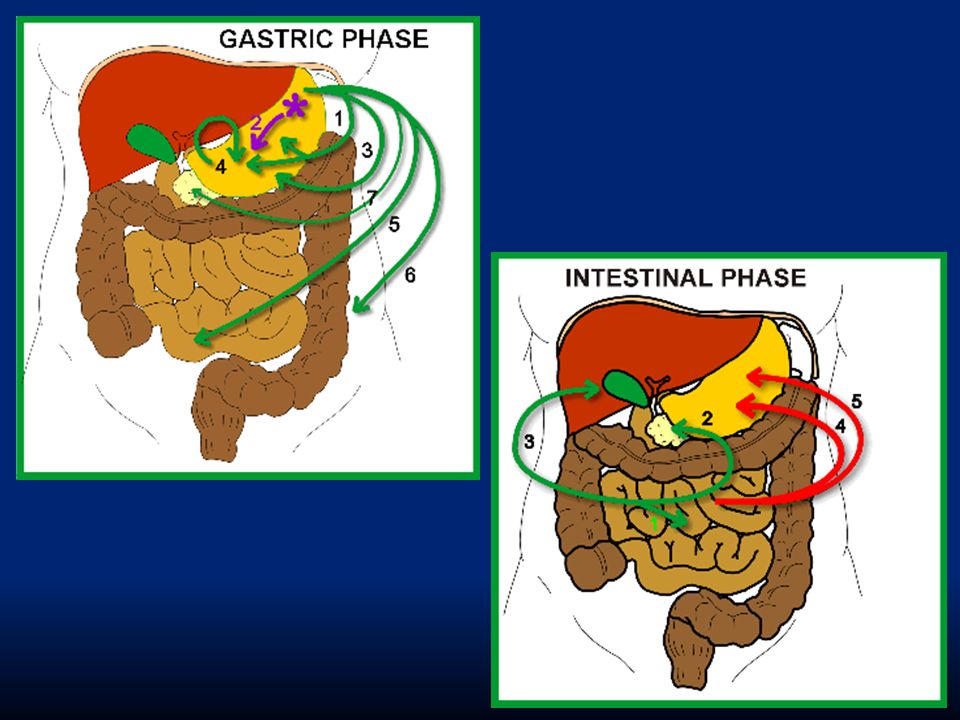
113
Fase gastrica: rende conto del 70% della secrezione gastrica e deriva dall’ingresso del cibo nello stomaco. Fa capo a riflessi vago-vagali e a riflessi locali che stimolano il rilascio di gastrina dalle cellule G. Fase intestinale: rende conto del rimanente 10% della secrezione. Dipende dalla presenza di cibo nel duodeno che evoca il rilascio di gastrina da cellule duodenali e che ha lo stesso effetto della gastrina “gastrica”.

Presentazioni simili